Cell Biology Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Discussion and Conclusion
In this study, we examined the growth of several bladder cells after EGF treatment, and we compared their responsiveness to EGF with the activation of MAPK and STAT which was induced by EGF. In our bladder cells, EGF induced the activation of MAPK as well as STAT. Moreover, EGF-activated STATs enhanced the expression of a CDK inhibitor, p21waf1. It is well known that activation of MAPK by EGFR-signaling and subsequent activation of several transcriptional factors enhances the cell proliferation in several cell types. On the other hand, Chin et al. reported that EGF-induced activation of STAT induced the expression of CDK inhibitor, p21waf1, and the expression of p21waf1 inhibited the growth of epidermal cancer, A431 (1996). In our bladder cell system, EGF activated the MAPK strongly in 1T-1 and A431, and slightly in 1T-2, 1T-3 and T24, but marginally in RT4. Futhermore, EGF activated STAT1 in 1T-3 and A431, and STAT1 and STAT3 in T24, and enhanced the expression of p21waf1 mRNA in T24, 1T-2, 1T-3, and A431. As we shown in Figure 6, EGF inhibited the growth of A431 and T24 cells, probably because STAT-mediated negative effect may supersede the positive effect mediated by the MAP kinase pathway. EGF markedly stimulated 1T-1 cells (and also 1T-2 and 1T-3 cells although less so), probably because the MAP kinase-mediated positive effect did not exert an inhibitory effect by the STAT pathway. In RT4 cells, EGF had no effect, probably because neither pathway was not effectively activated. All of our bladder cells expressed EGF receptor, and exogenous EGF enhanced the tyrosine-phosphorylation of the EGF receptor in the cells (Table I). However, the responsiveness to EGF did not correlate with the number of EGF receptors nor their affinity to the ligand.
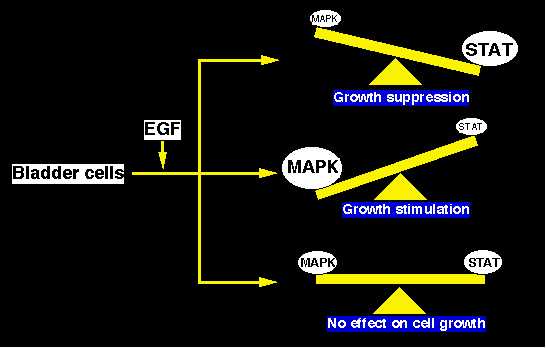 Figure 6
Figure 6It was reported that activation of STAT required the activation of tyrosine kinase of the JAK family (Darnell Jr. et al., 1997). However, David et al. (1996) reported that EGF- or amphiregulin-induced STAT activation required the intrinsic kinase activity of the EGF receptor but not the presence of JAK1. Leaman et al. (1996) also reported that JAK1, JAK2, and TYK2 was not required for STAT activation after EGF treatment, but kinase activity in EGF receptor was required, directly or indirectly. On the other hand, Wen et al. (1995) reported that maximum activation for STAT1 and STAT3 required not only tyrosine phosphorylation but also serine phosphorylation. They suggested that the phosphorylation of serine residue at 727 in STAT1 may be carried out by activated MAP kinase (Wen et al., 1995). Thus MAPK pathway and STAT pathway has an intimate crosstalk, and these pathway may not be activated independently. Therefore it might be difficult to evaluate MAPK pathway as a mitotic signal and STAT pathway as an inhibitory signal separately. As a simplified model system, in our bladder cells, the balance between the activated STAT-induced p21waf1 and MAP kinase activities is closely associated with the growth-responsiveness to EGF. Although we do not have any data accounting for the different regulation of the EGFR-signaling among our bladder cells at present, activation of STAT kinases and upper-stream regulators for MAPK, and protein phosphatase for STATs pathway and for MAPK pathway may be associated with different responsiveness to EGF.
Several investigators have examined the expression of EGF receptors, and their ligands in bladder tumors, and evaluated the malignancy of the tumor and attempted to predict the prognosis of the patients. The data presented here clearly demonstrates that growth-responsiveness of the bladder cancer cells and immortalized bladder cells to EGF is dependent on the activation of different signal transduction molecules but not on the number and affinity of EGF receptors. We are currently investigating the expression and activation of EGF receptors, the activation of MAPK and STAT, and the induction of p21waf1 in several human bladder cancer tissues. Then we are trying to evaluate the relationship of the activation of these signal transduction molecules with their clinical behavior and the prognosis of the patients.
| <= Results | DISCUSSION & CONCLUSSIONS | References => |
| Discussion Board | Next Page | Your Poster Session |