Invited Symposium: Na-H Exchangers and Intracellular pH Regulation
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Results
LITHIUM RESISTANCE IN DIFFERENT SOD2 MUTANTS.
The sod2 protein functions to remove internal sodium however it can also transport lithium ions in exchange for protons. This makes it possible to assess functional effects of different mutations of sod2 in vivo by measuring lithium tolerance in intact S. pombe. Since Li+ is much more toxic to cells than Na+, one can use relatively low LiCl concentrations that eliminates possible osmotic effects.
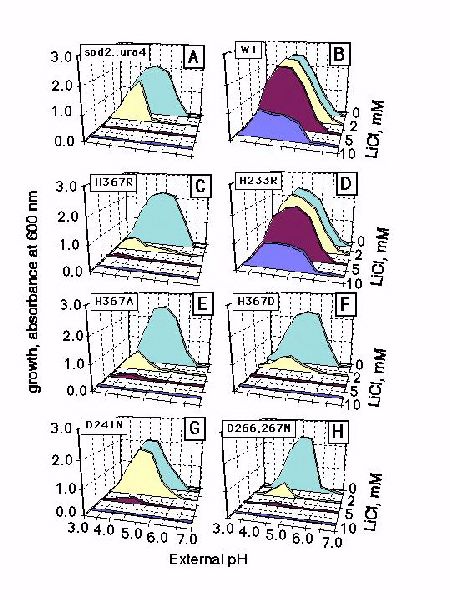 Fig. 2: LiCl tolerance in wild type and sod2 mutants at various external pHs. The ordinate represents growth (OD600 nm) in liquid minimal medium for 24 hours (A-F) or 48 hours (G-H), measured as described in "Materials and Methods". LiCl tolerance was measured in: disrupted sod2::ura4 mutant (A); the same mutant transformed with pWH5 bearing wild-type sod2 (B), and in a series of sod2 mutants introduced into sod2::ura4 with pWH5-sod2 derivatives carrying the respective mutations (C-H): H367R (C); H233R (D); H367A (E); H367D (F); D241N (G); D266,267N (H)..
Fig. 2: LiCl tolerance in wild type and sod2 mutants at various external pHs. The ordinate represents growth (OD600 nm) in liquid minimal medium for 24 hours (A-F) or 48 hours (G-H), measured as described in "Materials and Methods". LiCl tolerance was measured in: disrupted sod2::ura4 mutant (A); the same mutant transformed with pWH5 bearing wild-type sod2 (B), and in a series of sod2 mutants introduced into sod2::ura4 with pWH5-sod2 derivatives carrying the respective mutations (C-H): H367R (C); H233R (D); H367A (E); H367D (F); D241N (G); D266,267N (H)..
Figure 2 (panel A) confirms that the sod2::ura4 disruption mutant is highly sensitive to external lithium. LiCl at a concentration exceeding 2 mM, completely arrests growth in the pH range of 3.5 to 6.5. Transformation of the sod2::ura4 strain with the plasmid carrying wild-type sod2, restores resistance to as much as 10 mM of LiCl within the same pH range (Fig. 2B). The H367R mutation of sod2 affects the restoration of LiCl resistance markedly. It is as sensitive to LiCl as the sod2::ura4 disruption mutant (Fig. 2C). Replacements of the other seven His residues of sod2 with Arg did not change lithium tolerance in transformant cells. An example of such a 'neutral' His substitution (H233R mutant) is shown in Fig. 1D. There was no difference in sensitivity to LiCl between this mutant and cells transformed with the wild-type sod2 plasmid. Replacement with either Ala (Fig. 2E) or an Asp (Fig. 2F) followed by transformation into the sod2::ura4 strain resulted in cells that were sensitive to external Li+ at all pH's. In the next series of experiments, three Asp residues located in putative transmembrane segments of the sod2 protein, were substituted with the uncharged analog Asn. In the bacterial antiporter the same change in aspartate residues within this conserved '3D-motif' inactivate the protein. When sod2 was mutated and transformed into the sod2::ura4 strain, lithium tolerance was not restored in cells with either the D241N (Fig. 2G) or the D266,267N (Fig. 2H) mutation.
EFFECT OF EXTERNAL pH ON INTERNAL pH.
Because the proton gradient acts as the sole driving force for Na+ or Li+ extrusion mediated by sod2 we examined the effect of external pH on internal pH in the sod2::ura4 strain transformed with the wild type sod2. We then determined the DpH on the membrane. The results are shown in Figure 3. At acidic pHs the DpH is clearly the largest. The proton gradient declines progressively with alkalinization of the medium and at external pHs above 6 the DpH becomes minimal while above pH 6.5 gradient is reversed.
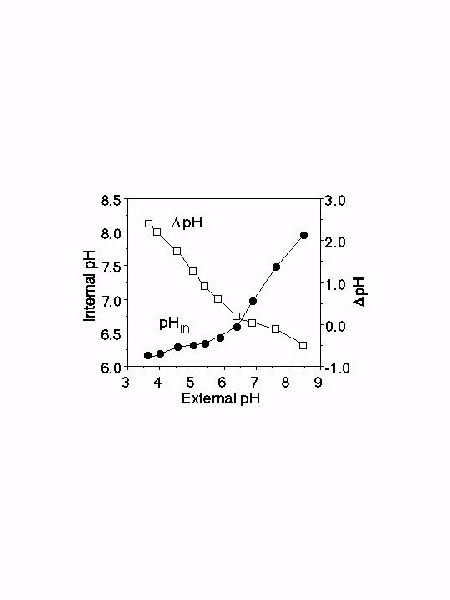 Fig. 3: pH homeostasis and DpH in S. pombe sod2::ura4 transformed with pWH5-sod2. Intracellular pH values at different pHs of the medium ( ) were measured as described. Each point represents the average of two independent measurements. The standard deviation was less than 6%. The DpH on the plasma membrane ( ) was calculated by subtraction of intracellular pH values.
Fig. 3: pH homeostasis and DpH in S. pombe sod2::ura4 transformed with pWH5-sod2. Intracellular pH values at different pHs of the medium ( ) were measured as described. Each point represents the average of two independent measurements. The standard deviation was less than 6%. The DpH on the plasma membrane ( ) was calculated by subtraction of intracellular pH values.
EXPRESSION OF sod2 MUTANTS. We examined expression of the various sod2 mutants to confirm that they were expressed and correctly targeted. We added the highly immunoreactive hemagglutinin (HA) tag to the carboxyl terminal of the protein and then examined expression under control of the endogenous sod2 promoter. The tag was added to the control and to sod2 mutants that exhibited significant changes in phenotype (wild type; H367R, H367A, H367D; D241N, and D266,267N). We used monoclonal anti-HA antibody to examine the level of the sod2 mutant proteins in membrane preparations from transformants of sod2::ura4 (Fig. 4).
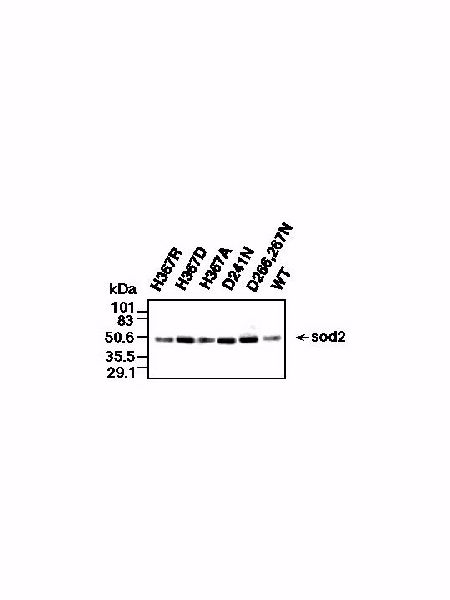 Fig. 4: Immunodetection of sod2 protein in S. pombe membranes. Crude membranes were solubilized with dodecyl maltoside and analyzed using SDS-PAGE followed by immunoblotting with anti-HA.
Fig. 4: Immunodetection of sod2 protein in S. pombe membranes. Crude membranes were solubilized with dodecyl maltoside and analyzed using SDS-PAGE followed by immunoblotting with anti-HA.
In all analyzed cases introduced mutations did not prevent either synthesis of sod2 or its membrane targeting. Therefore, differences in lithium tolerance among the mutant strains could not be attributed to an inability to express the sod2 mutant proteins.
ACTIVE 22Na EFFLUX FROM THE WHOLE CELLS.
To examine sod2 function directly in various mutants we measured 22Na transport in the yeast strains. It was necessary to examine transport in acidic medium where the cells can maintain a significant DpH on the plasma membrane (Fig. 3). Cells were pre-loaded with 22Na at pH 7.0, briefly washed, and quickly transferred into isotope-free medium of pH 4.0. Sodium efflux was assessed in the sod2::ura4 strain transformed with either wild type or mutant sod2 genes.
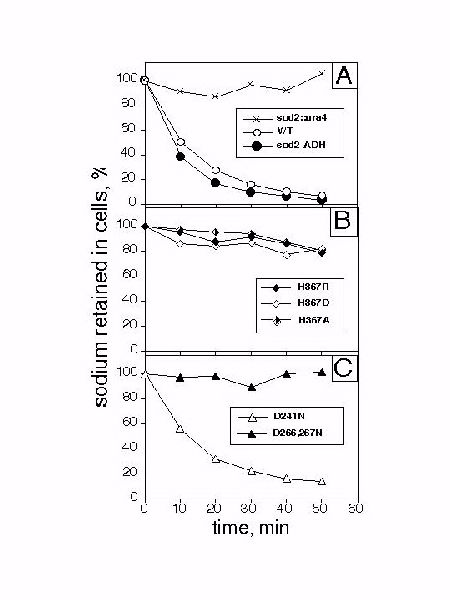 Fig. 5: Active sodium transport in intact S. pombe. Cells were equilibrated with 22Na+ (0.5 mCi/ml) for 1 hour in the presence of 10 mM unlabeled NaCl at pH 7.0. 22Na+ transport at pH 4.0 was monitored by filtration after dilution into 22Na+-free medium as described in the "Materials and Methods". Radioactive Na+ remaining in the cells is expressed relative to that recorded at T=0 minutes.
Fig. 5: Active sodium transport in intact S. pombe. Cells were equilibrated with 22Na+ (0.5 mCi/ml) for 1 hour in the presence of 10 mM unlabeled NaCl at pH 7.0. 22Na+ transport at pH 4.0 was monitored by filtration after dilution into 22Na+-free medium as described in the "Materials and Methods". Radioactive Na+ remaining in the cells is expressed relative to that recorded at T=0 minutes.
Initial results (Fig. 5A) confirmed that Na+ efflux was absent in disrupted strain while sod2-overexpressing cells demonstrated greater sodium efflux than the wild type (Fig. 5A). This data confirms that 22Na efflux reflects sod2 function. When we examined the sod2::ura4 strain containing the H367R, H367A and H367D mutants, we found only residual 22Na export (Fig. 5B). Similarly, the D226,227N mutant failed to export 22Na (Fig. 5C). In contrast, the D241N containing strain demonstrated rapid efflux of 22Na. This result was surprising considering that growth of sod2::ura4 containing this mutant was inhibited by the addition of lithium (Fig. 2G).
Na+ DEPENDENT H+ UPTAKE.
Sod2 catalyzes coupled transmembrane movements of sodium ions and protons. Therefore, it is possible to measure sod2 activity by measuring proton influx that is driven by a high internal sodium gradient. We used this approach to measure the activity of sod2 at an external pH of 6.1. At this external pH S. pombe cells show a reduced rate of growth and the DpH on the plasma membrane is small, less than 0.5 pH units (Fig. 3). Therefore it is relatively easy to load these cells with considerable amounts of Na+. We created an artificial DNa (high Na+ concentration inside the cell) by incubation of cells in medium with 0.1 M NaCl at pH 6.1 and subsequently transferring the cells into the same buffer devoid of NaCl.
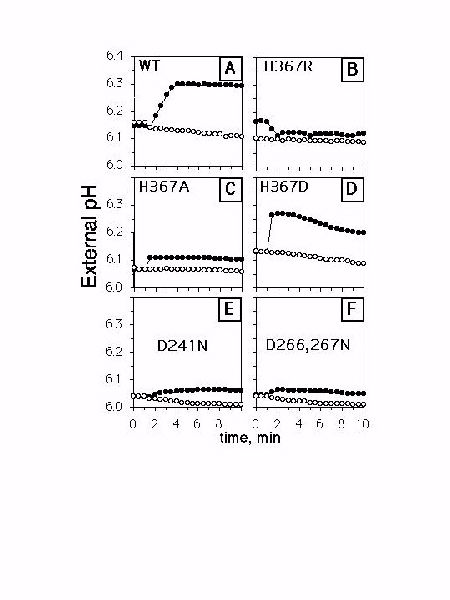 Fig. 6: Na+ dependent H+ fluxes mediated by S. pombe. Cells carrying either the wild-type or mutant sod2 were either loaded with 100 mM NaCl (filled symbols) or not (open symbols). They were then briefly washed and added to the experimental buffer; H+ influx was then monitored as described in "Materials and Methods". Results are typical of at least three separate experiments. (A) wild type; (B) H367R; (C) H367A; (D) H367D; (E) D241N; (F) D266,267N.
Fig. 6: Na+ dependent H+ fluxes mediated by S. pombe. Cells carrying either the wild-type or mutant sod2 were either loaded with 100 mM NaCl (filled symbols) or not (open symbols). They were then briefly washed and added to the experimental buffer; H+ influx was then monitored as described in "Materials and Methods". Results are typical of at least three separate experiments. (A) wild type; (B) H367R; (C) H367A; (D) H367D; (E) D241N; (F) D266,267N.
By monitoring the pH of a weakly buffered external medium, we found that within a few minutes, cells containing the wild type sod2 exhibit Na+ dependent proton uptake (Fig. 6A, filled symbols). Cells that were not loaded with sodium did not alkalinize the external medium (Fig. 6A, empty symbols). In cells bearing sod2 with the H367R (Fig. 6B) or the H367A (Fig. 6C) mutation we found no alkalinization of the external medium. However, a rapid proton flux was observed in the H367D mutant cells preloaded with NaCl (Fig. 6D). The magnitude of resulting pH shift was comparable to that produced by wild type cells. In addition, the H367D mutant alkalinized the experimental medium much faster than the wild type. Both '3-D motif' AspĆAsn mutants were not capable of taking up protons in exchange for Na+ at the external pH of 6.1 (Fig. 6E, F).
| <= Materials & Methods | RESULTS | Discussion & Conclussions => |
| Discussion Board | Next Page | Your Symposium |