Invited Symposium: Behaviour-Induced Neural Events after Brain Injury
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
Recent work has suggested that motor learning, but not mere motor activity, is associated with significant morphological changes (addition of synapses, dendrite outgrowth) in brain areas known to participate in the control of movement: cerebellar and motor cortex (Black et al., 1990; Kleim et al., 1996, 1997). Early postnatal exposure to alcohol during the "brain growth spurt" (first 10 postnatal days in rats) results in deficits in motor performance (Goodlett et al., 1991; Melcer et al., 1995) and in permanent loss of cerebellar neurons (e.g. Marcussen et al., 1994; Pauli et al., 1995). Previously we demonstrated that complex motor skill learning significantly improves ("rehabilitates") motor performance in animals exposed to ethanol postnatally and that it causes synaptogenesis in cerebellar cortex damaged by exposure to neonatal binge-like ethanol (Klintsova et al., 1997). The present study investigated how neonatal alcohol exposure affects the neurons in motor cortex and if motor skill learning has similar rehabilitative effects on this brain area.
Materials and Methods
Subjects
A total of 130 rats (65 female, 65 male) from 17 litters resulting from timed pregnancies of adult Long-Evans rats (Simonsen Labs, Gilroy, CA) bred in the Indiana University-Purdue University, Indianapolis (IUPUI) vivarium were used in this study. See complete description of the Material and Methods in Klintsova et al. (1998), which used the same animals.
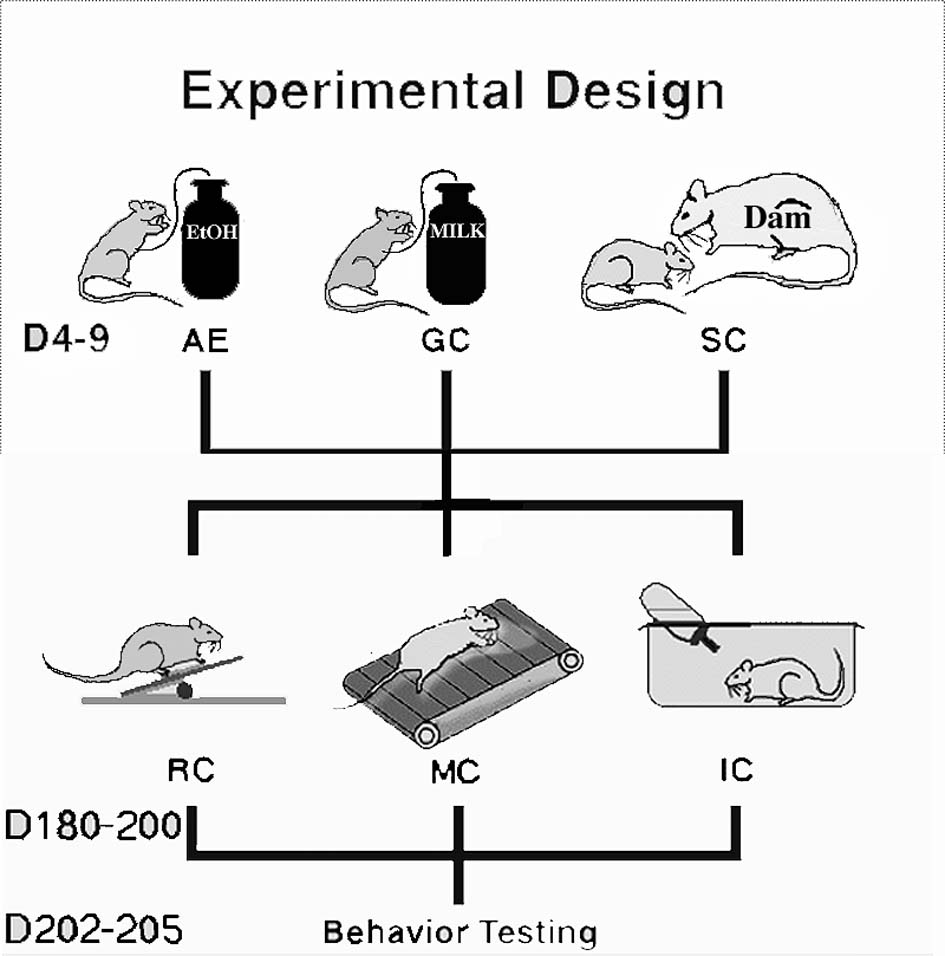
Artificial Rearing and Alcohol ExposureOn PD 4, pups were assigned randomly within litter and sex to three groups (Fig. 1):
 Click to enlarge
Fig. 1: Design of the study: pups were evenly distributed across three postnatal treatment groups (AE, GC and SC). When the animals were 6 month old, they spent 20 days in one of three conditions: inactive (IC), motor (MC) or rehabilitation (RC).
Click to enlarge
Fig. 1: Design of the study: pups were evenly distributed across three postnatal treatment groups (AE, GC and SC). When the animals were 6 month old, they spent 20 days in one of three conditions: inactive (IC), motor (MC) or rehabilitation (RC).
Alcohol Exposed (AE) - artificially reared pups given a 10.2% (v/v) ethanol solution in milk formula on two consecutive feedings each day (4.5 g/kg/day) from PD 4-9; gastrostomy control (GC) - artificially reared pups given matched isocaloric maltose/dextrin solutions on PD 4-9; suckle controls (SC) - reared normally by lactating dams (Fig.1). The rats assigned to the artificial rearing groups were surgically implanted with intragastric feeding tubes under methoxyflurane (Pitman-Moore, Mundelein, IL) anesthesia on PD 4 and reared using well-established procedures.
All intragastric feedings used a customized milk formula delivered every two hours. Alcohol (or maltose/dextrin) was provided on the first two feedings after 8:00 a.m. each morning; all other feedings used milk formula alone. Each day, the rats were fed a volume of formula equal to 33% of the mean body weight (in g) of the litter being reared. Seventy minutes after the end of the second alcohol feeding on PD 6, a 20-microliter sample of blood was collected from a tail-clip of each artificially reared pup. The blood from the alcohol-treated pups was assayed with a enzymatic assay for ethanol content (Sigma kit #332-BT, St. Louis, MO).
Artificially reared offspring were fostered back to lactating dams on PD 12; all rats were weaned at 25 days of age, and housed 2-4 per cage with same-sex littermates thereafter.
Rehabilitative Motor Skill Training Procedure
Rats were transferred to the University of Illinois vivarium at age 60-100 days. They were housed in same-sex pairs until they reached age 180 days. At that point, animals from each treatment group (SC, GC and AE) were assigned to one of three training conditions (Fig. 1): an inactive condition (IC), a motor control condition (MC) or a rehabilitative condition (RC). RC rats were given 5 trials per day on an elevated obstacle course for 20 days as described previously [Black et al., 1990; Kleim et al., 1996]. The obstacles included a horizontally placed wooden ladder, narrow rods, a link chain, barriers on narrow beams, a rope ladder, an elastic cord, ascending and descending stairs, and a narrow v-shaped metal bridge. Each RC animal was pair matched with an animal in the motor control condition. MC rats were forced to traverse a flat, opaque, oval Plexiglas track equal in length to the acrobatic course. IC animals were housed individually and received no motor training or extra-cage motor activity but were handled for about 5 min. each day. At the completion of the 20 days of rehabilitative motor skill training, the motor ability of each rat was tested on parallel bars, a rotating rod and a vertically suspended rope. Behavioral data was analyzed using the SAS general linear model procedure and were conducted at a significance level of alpha = 0.05. Two-way repeated measures ANOVA with GROUP (postnatal treatment: SC, GC or AE) and TRAINING (IC, MC or RC) as between-group factors was used to analyze the results of all three behavioral tests. The Student Newman-Keuls post-hoc (alpha = 0.05) test was used for a posteriori comparisons among individual groups.
Golgi Impregnation ProcedureAfter completion of testing animals were anesthetized with sodium pentobarbital and perfused with 0.9% NaCl (pH 7.3). Brains were placed in 100 ml of Golgi-Cox solution for 5 weeks, followed by 2 days in 30% sucrose solution (Glaser & Van der Loos, 1981). Coronal 150 �m sections were taken from primary motor cortex, mounted on slides, developed in ammonium hydroxide and coverslipped. Pyramidal neurons in layer II/III were traced with the aid of a camera lucida (730x). The apical and basal dendritic arborization of pyramidal neurons was analyzed by counting the number of intersections between dendrites and an overlay of concentric rings with 10 �m inter-ring distance (Sholl, 1956).
Tissue preparation for Electron Microscopy and Stereological Methods
A subgroup of the animals was used for EM study immediately after completion of 20 days complex motor task training or control procedures. Animals were anesthetized with sodium pentobarbital and perfused with mixture of 2% glutaraldehyde/2% paraformaldehyde. Coronal sections through the M1 cortical area were postfixed, dehydrated and embedded in Epon. One-micron sections were used to estimate neuronal density in layer II/III using unbiased stereology ("physical disector"). Series of 12-15 ultrathin sections were used for estimation of synaptic density in neuropile of layer II/III using physical disector as well.
Results
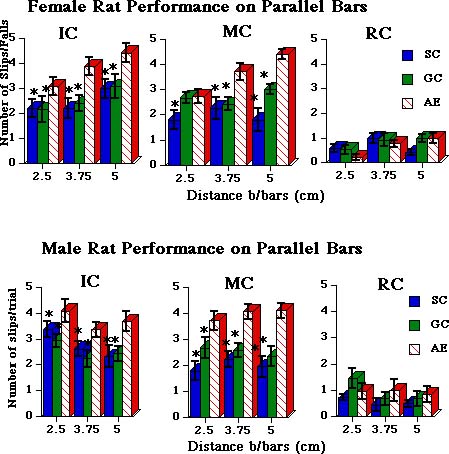
All animals, including AE, significantly improved their motor skills after 20 days of complex motor training. Results of the tests were reported previously in Klintsova et al.(1998) (Fig.2).
 Click to enlarge
Fig. 2: Results of female (top) and male (bottom) rats� performance on parallel bars. Note that both female and male AE rats demonstrate a significantly higher number of slips/falls than their SC and GC littermates after 20 days in IC or MC. AE-RC rats are no longer different from controls (right graphs).
Click to enlarge
Fig. 2: Results of female (top) and male (bottom) rats� performance on parallel bars. Note that both female and male AE rats demonstrate a significantly higher number of slips/falls than their SC and GC littermates after 20 days in IC or MC. AE-RC rats are no longer different from controls (right graphs).
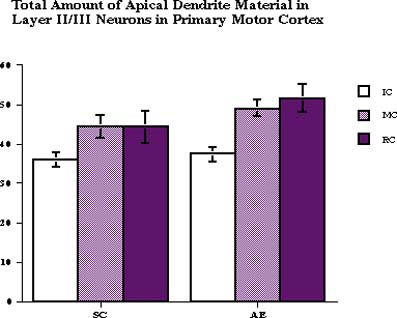
A 2-way ANOVA with TREATMENT and CONDITION as between-subject factors revealed the effect of adult CONDITION (F2,35=3.34, p<0.05) on the total amount of apical dendrite material: there were more dendritic intersections with concentric rings after MC and RC (Fig. 3).
 Click to enlarge
Fig. 3: The total number of intersections between apical dendrites of layer II/III pyramidal neurons and concentric ring overlay. Both motor activity and motor learning had an effect on total amount of apical dendrite material.
Click to enlarge
Fig. 3: The total number of intersections between apical dendrites of layer II/III pyramidal neurons and concentric ring overlay. Both motor activity and motor learning had an effect on total amount of apical dendrite material.
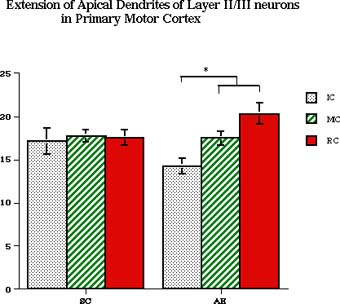
There was no interaction between these factors. Neonatal TREATMENT and adult CONDITION had an effect on the extention of apical dendrite in AE, but not SC animals (Fig. 4): within AE animals there was a decreased length of apical dendrite in AE-IC animals and consecutive increase of its length in MC and RC rats (p<0.05).
 Click to enlarge
Fig. 4: The extention of apical dendrite in layer II/III pyramidal neurons: there are shorter apical dendrites in AE-IC animals and significant increase of apical dendrite extention after 20 days of rehabilitation.
Click to enlarge
Fig. 4: The extention of apical dendrite in layer II/III pyramidal neurons: there are shorter apical dendrites in AE-IC animals and significant increase of apical dendrite extention after 20 days of rehabilitation.
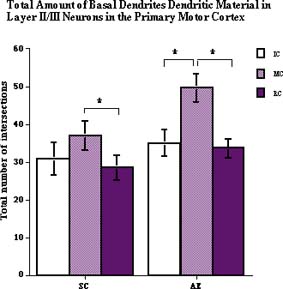
Adult CONDITION had a significant effect on total amount of basal dendritic material (F2,35=2.86, p<0.05): there was significantly more dendrite intersections with concentric circles in layer II/III pyramidal neurons of both SC and AE animals after running on the track (Fig.5).
 Click to enlarge
Fig. 5: The total number of intersections between basal dendrites of layer II/III pyramidal neurons and concentric ring overlay.Only motor activity had an effect on total amount of basal dendrite material.
Click to enlarge
Fig. 5: The total number of intersections between basal dendrites of layer II/III pyramidal neurons and concentric ring overlay.Only motor activity had an effect on total amount of basal dendrite material.
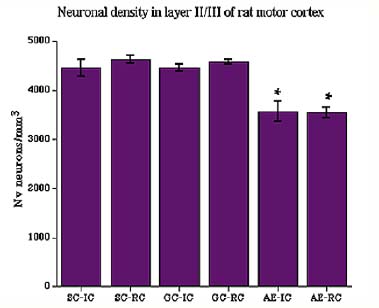
Analysis of neuronal density in layer II/III with 2-way ANOVA demonstrated a significant effect of postnatal tretament (F2,36=38.58, p<0.01): density of neurons was significantly lower in both AE-IC and AE-RC groups (Fig.6).
 Click to enlarge
Fig. 6: Neuronal density in layer II/III of motor cortex. Animals exposed to ethanol during early postnatal period had decreased density of neurons (AE-IC and AE-RC).
Click to enlarge
Fig. 6: Neuronal density in layer II/III of motor cortex. Animals exposed to ethanol during early postnatal period had decreased density of neurons (AE-IC and AE-RC).
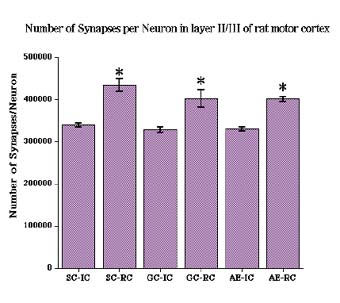
Analysis of the number of synapses/neuron in the neuropile of layer II/III revealed an effect of adult CONDITION (F2,36=106.23, p<0.001) (Fig. 7). Subsequent multiple comparisons (Student Neuman-Keuls, p<0.05) revealed that animals in all three postnatal groups had significantly more synapses per neuron after 20 days of complex motor learning.
 Click to enlarge
Fig. 7: Number of synapses per neuron in layer II/III was significantly increased after 20 days of complex motor task learning.
Click to enlarge
Fig. 7: Number of synapses per neuron in layer II/III was significantly increased after 20 days of complex motor task learning.
Discussion and Conclusion
This study is a logical continuation of a project searching for possibilities to remediate behavioral and morphological damage resulted from developmental exposure to alcohol. As we demonstrated previously, animals exposed to alcohol in a binge-like manner during an early postnatal period (a model of the third trimester of human pregnancy) can benefit from learning a complex motor task (Klintsova et al., 1997, 1998): after 10 or 20 days of learning the task alcohol-exposed animals were as succesful as control animals in specific motor tests and had an increased number of parallel fiber-Purkinje cell connections in cerebellar cortex.
The effect of developmental exposure to ethanol depends on the time of exposure: there is a suggestion that the window of greatest vulnerability for the neocortex occurs within the prenatal period of neurogenesis (Mooney et al., 1996). Postnatal alcohol exposure did not result in changes in total number of large pyramids in layer V of the neocortex (Mooney et al., 1996), although there is some evidence for prenatal alcohol-induced cell loss in certain cortical subdivisions, such as somatosensory cortex (Miller, 1996; Miller & Potempa, 1990).
Here we demonstrate that postnatal exposure to alcohol results in changes to the dendritic tree of pyramidal neurons in layer II/III and in overall decreased density of neurons in this layer of M1. Interestingly, apical and basal dendrites of pyramidal neurons are affected differently by alcohol and later - by adult experience: apical dendrites, receiving predominantly parallel afferents from other cortical areas, are shorter in AE animals and significantly more extended after rehabilitative complex motor skill training. Quite the opposite, the major target of thalamic afferents - basal dendrites - are not affected by exposure to ethanol, but their dendritic tree is significantly increased after 20 days of running on the flat track.
Decreased density of neurons in the M1 area may indicate increased neuronal death after ethanol exposure: an evaluation of the total number of neurons in this area of SC and AE animals is under way.
The increased number of synapses per neuron in layer II/III demonstrates synaptogenesis in response to complex motor training in adult motor cortex of animals exposed to ethanol postnatally. These results indicate that exposure to binge-like alcohol in the early postnatal period results in neuronal damage in different brain areas involved in motor activity and control and that complex motor skill learning can provide a therapeutic effect on some of this damage (motor performance and neuronal plasticity).
References
- Black JE, Isaacs KR, Anderson BJ, Alcantara AA and Greenough WT. (1990) Learning causes synaptogenesis, whereas motor activity causes angiogenesis, in cerebellar cortex of adult rats. Proc. Natl. Acad. Sci., 87, 5568-5572.
- Glaser EM, and Van der Loos H. (1981) Analysis of thick brain sections by obverse-reverse computer microscopy: Application of a new, high clarity Golgi-Nissl stain. J Neurosci Methods, 4, 117-125.
- Goodlett CR, Thomas JD and West JR. (1991) Long-term deficits in cerebellar growth and rotarod performance of rats following "binge-like" alcohol exposure during the neonatal brain growth spurt. Neurotox. Teratol., 13, 69-74.
- Kleim JA, Lussnig E, Schwarz ER, Comery TA and Greenough WT. Synaptogenesis and FOS expression in the motor cortex of the adult rat after motor skill learning. J Neurosci, 16 (1996), 4529-4535.
- Kleim JA, Vij K, Ballard DH and Greenough WT. (1997) Learning-dependent synaptic modification in the cerebellar cortex of the adult rat persist for at least four weeks. J Neurosci, 17(2), 717-721.
- Klintsova AY, Matthews J T, Goodlett CR, Napper RMA and Greenough WT. (1997) Therapeutic motor training increases parallel fiber synapse number per Purkinje neuron in cerebellar cortex of rats given postnatal binge alcohol exposure. Alcoholism: Clinical and Experimental Research, 21:1257-1263.
- Klintsova AY, Cowell RM, Swain RA, Napper RMA, Goodlett CR, and Greenough WT. (1998) Therapeutic effects of complex motor training on motor performance deficits induced by neonatal binge-like alcohol exposure inrats. I. Behavioral results. Brain Res, 800, 48-61.
- Marcussen BL, Goodlett CR, Mahoney JC, and West JR. (1994). Developing rat Purkinje cells are more vulnerable to alcohol-induced depletion during differentiation than during neurogenesis. Alcohol, 11, 147-156.
- Melcer T, Gonzalez D, Somes C, Riley EP. (1995) Neonatal alcohol exposure and early development of motor skills in alcohol preferring and nonpreferring rats. Neurotoxicol and Teratol, 17, 103-110.
- Miller MW. (1996). Limited ethanol exposure selectively alters the proliferation of precursors cells in the cerebral cortex. Alcoholism: Clinical and Experimental Research, 20:139-143.
- Miller MW and Potempa G. (1990) Numbers of neurons and glia in mature rat somatosensory cortex: effects of prenatal exposure to ethanol. J. Comp. Neurol., 293, 92-102.
- Mooney SM, Napper RMA and West JR. Long-term effect of postnatal alcohol exposure on the number of cells in the neocortex of the rat: A stereological study. Alcoholism Clin. Exp. Res., 20, 615-623, 1996.
- Pauli J, Wilce P, Bedi KS. (1995) Acute exposure to alcohol during early postnatal life causes a deficit in the total number of cerebellar Purkinje cells in the rat. J Comp Neurol, 360:506-512.
- Sholl DA. (1956) The organization of the cerebral cortex. London: Methuen.
| Discussion Board | Previous Page | Your Symposium |