Pharmacology & Toxicology Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
13,14-dihydro, 15-keto-PGD2(DK), which is a stable metabolite of PGD2, produces a sharp reduction of Isc on stimulated canine colonic epithelium. This effect is noted with a variety of agonists including carbachol (Carb), cyclopiazonic acid (CPA), histamine, and PGE2. It has been our observation that this effect appears to be much more pronounced with agonists that are believed to exert their effects via changes in intracellular Ca2+ (i.e. carbachol and CPA), than with agonists that work via cAMP (e.g. forskolin). With forskolin (FSK) the addition of DK, often produces little or no change in Isc. Using a group of selected agonists (Carb, CPA, and FSK), the present study seeks to further investigate the role of external Ca2+ in mediating changes in Isc, and what role if any DK plays in this process. Our objective is therefore to answer the following questions:
- Is there a relationship between agonist-mediated increases in Isc and external Ca2+ (in particular serosal Ca2+)?
- Can the effects of DK be explained by demonstrating an interaction with either internal or external Ca2+, i.e. does DK appear to interfere in some way with normal Ca2+ dynamics?
Materials and Methods
Many of the procedures used in the present study were described in detail in our earlier publications (Rangachari et al., 1992). Briefly, adult dogs of either sex were euthanised with sodium pentobarbital (100 mg/kg) and the proximal colon quickly excised and immersed in oxygenated Krebs solution warmed to 37. The tissues were bathed on both sides with warm (37) "Ca2+-free" Agonist Concentration-Response Curve's
Briefly, multiple pieces of tissue from the same dog were set-up in normal Kreb's buffer and allowed to equilibrate for approx. 30 mins before beginning the experiment. Control tissues were washed 3x with normal Kreb's on both the luminal and serosal sides of the tissue. "Ca-free" tissues were established by washing (3x) the serosal side of the tissue with Ca-free Kreb's (no Ca2+ and 1 Ca2+ Concentration-Response Studies
Tissues were set-up in "Ca-free" conditions as described above. After a period of 10-15 min, cumulative concentration-response curves were then constructed by adding increasing concentrations of Ca2+ back to the serosal bathing solutions. Several series of experiments were performed. Curves were constructed both in the presence and absence of a stimulating agonist (CPA), with the former giving the best responses. The following sequence was used in these experiments. After 10 min in Ca-free buffer the agonist (CPA or Carb) was added at a concentration of 10.
Results
 |
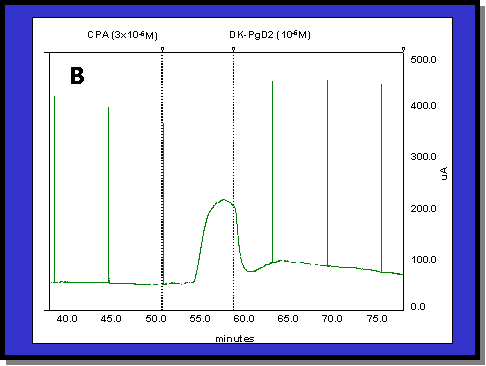 |
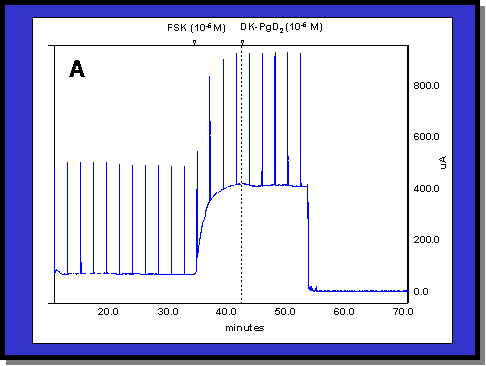
Fig. 1: (A) Typical colonic response to FSK (1 mM). The addition of 1 mM FSK produces a 200 mA/cm2 increase in Isc. The addition of DK (1 mM) has little or no effect on the Isc generated by FSK. (B) Typical colonic response to CPA (3 mM). The addition of 3 mM CPA produces a 90 mA/cm2 increase in Isc which is abruptly reversed by the addition of DK (1 mM)
.
 |
 |
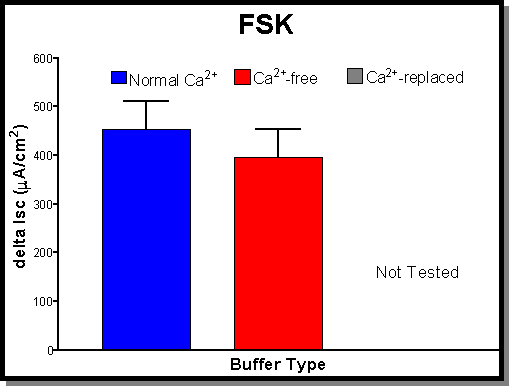 |
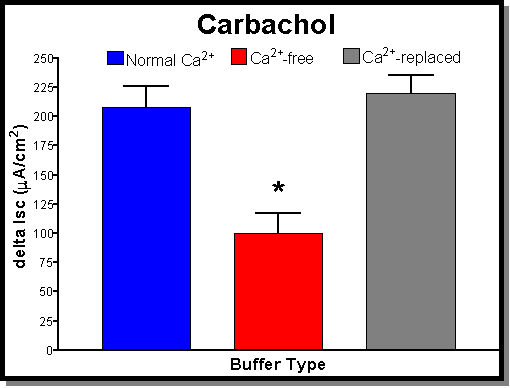
Fig. 2: (A) The addition of carbachol (100 m
M) on average produces an Isc of 208�
18 mA/cm2 (blue bar). Under Ca-free conditions (red bar) there is a significant drop in the carbachol response to a 100�17 mA/cm2 . This effect is reversible with the addition of Ca2+ (1.5 mM-normal Ca2+). Ca2+-replaced tissues (grey bar) had responses (220�
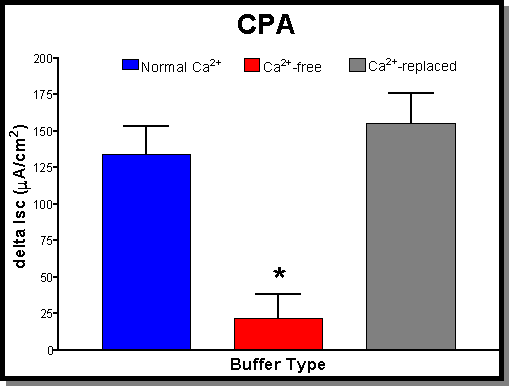
15 mA/cm2) that were not significantly different from controls. (B) The addition of CPA (30 mM) on average produces an Isc of 134�19 mA/cm2 (blue bar). Under Ca-free conditions (red bar) there is a significant drop in the CPA response to a 22�16 mA/cm2 . Once again this effect is reversible with the addition of Ca2+ (1.5 mM-normal Ca2+). Ca2+-replaced tissues (grey bar) had responses (155�21 mA/cm2) that were actually larger than control values. This difference was however not significantly different from controls. (C) The addition of FSK (30 mM) on average produces an Isc of 453�59 mA/cm2 (blue bar). Under Ca-free conditions (red bar) there is a small drop in the FSK response to a 397�57 mA/cm2 which was not statistically significant. The addition of normal Ca2+ (1.5 mM) was not tested with this agonist since there was no significant effect produced by removing Ca2+
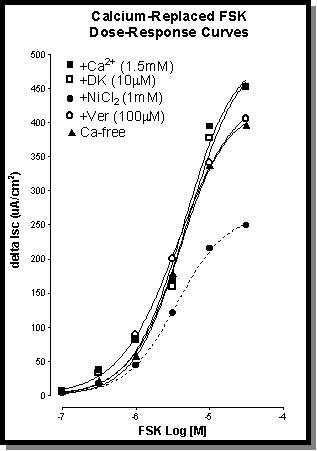 Fig. 3: Concentration response curves to FSK (0.1-30 m
M) showing the effects of pre-treatment with Ni2+ (!
-1mM), verapamil ("
-100 m
M), DK (0
-10 m
M) and Ca-free (_
) conditions. Only the addition of Ni2+ has an effect on the FSK responses.
Fig. 3: Concentration response curves to FSK (0.1-30 m
M) showing the effects of pre-treatment with Ni2+ (!
-1mM), verapamil ("
-100 m
M), DK (0
-10 m
M) and Ca-free (_
) conditions. Only the addition of Ni2+ has an effect on the FSK responses.
 |
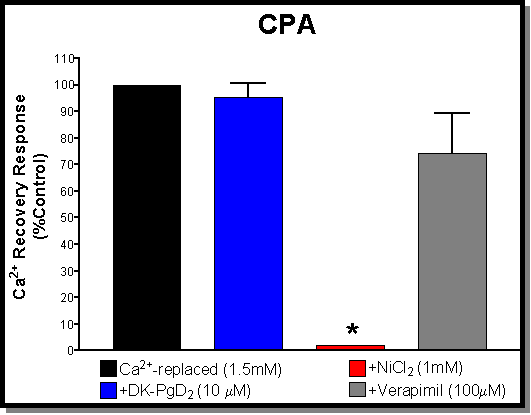 |
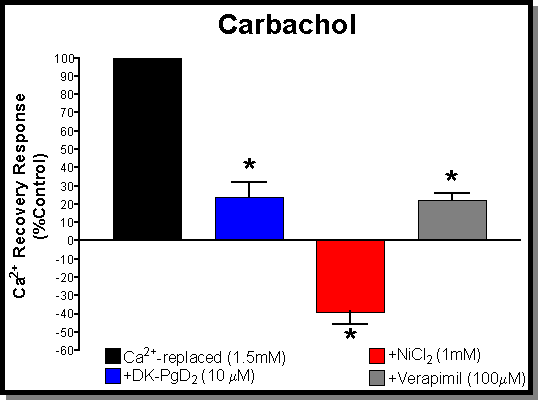
Fig. 4: (A) Antagonism of the "Ca2+-reversal" effect using carbachol (100 mM) as the agonist. All responses are normalised to control values obtained with carbachol on tissues that had serosal Ca2+ restored to normal levels (black bar-1.5mM). The other bars show the effects of pre-treatment with Ni2+ (red bar-1mM), verapamil (grey bar-100 mM), and DK (blue bar-10 mM) prior to restoring serosal Ca2+ levels. Both verapamil and DK caused a significant inhibition of the Ca2+-reversal effect. By far Ni2+ has the largest effect producing an inhibition that is greater than that achieved with Ca2+-free serosal buffer (the zero reference point). (B) Antagonism of the "Ca2+-reversal" effect using CPA (10 mM) as the agonist. All responses are normalised to control values obtained with CPA on tissues that had serosal Ca2+ restored to normal levels (black bar-1.5mM). The other bars show the effects of pre-treatment with Ni2+ (red bar-1mM), verapamil (grey bar-100 mM), and DK (blue bar-10 mM) prior to restoring serosal Ca2+ levels. In contrast to the results obtained with carbachol, both verapamil and DK have no significant effects on the Ca2+-reversal effect. Again Ni2+ had a large effect producing an inhibition similar to that achieved with Ca2+-free serosal buffer (both essentially abolish the responses to CPA)
.
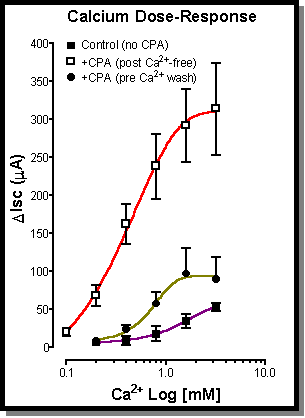 Fig. 5: The effect of CPA pre-treatment on Ca2+ concentration-effect curves. Under control conditions adding Ca2+ back to the serosal bathing solutions of Ca2+-free tissues results in a weak concentration-effect curve (#). Pre-treatment of the tissues with CPA (100 mM) prior to adding the Ca2+ results in a large potentiation of the Ca2+ concentration-effect curve as can be seen in the graph above (:). If the CPA is however added prior to the Ca2+-free washes it is a much less effective (!). It would appear that the CPA or a factor produced by the CPA that results in the potentiation of Ca2+ influx is removed by the washing protocol
Fig. 5: The effect of CPA pre-treatment on Ca2+ concentration-effect curves. Under control conditions adding Ca2+ back to the serosal bathing solutions of Ca2+-free tissues results in a weak concentration-effect curve (#). Pre-treatment of the tissues with CPA (100 mM) prior to adding the Ca2+ results in a large potentiation of the Ca2+ concentration-effect curve as can be seen in the graph above (:). If the CPA is however added prior to the Ca2+-free washes it is a much less effective (!). It would appear that the CPA or a factor produced by the CPA that results in the potentiation of Ca2+ influx is removed by the washing protocol
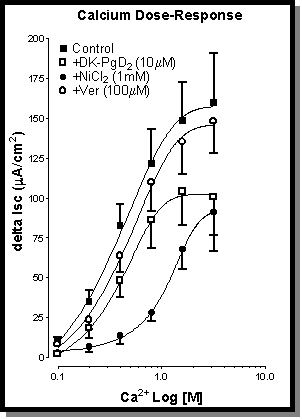 Fig. 6: CPA-stimulated Ca2+-concentration response curves showing the effects of pre-treatment with Ni2+ (!-1mM), verapamil ("-100 mM), and DK (0-10 mM). The addition of verapamil had no significant effect on control Ca2+ responses (#). Both DK and Ni2+ resulted in a significant inhibition of Ca2+ responses at all concentrations. Once again the Ni2+ was clearly resulted in the most effective inhibition
Fig. 6: CPA-stimulated Ca2+-concentration response curves showing the effects of pre-treatment with Ni2+ (!-1mM), verapamil ("-100 mM), and DK (0-10 mM). The addition of verapamil had no significant effect on control Ca2+ responses (#). Both DK and Ni2+ resulted in a significant inhibition of Ca2+ responses at all concentrations. Once again the Ni2+ was clearly resulted in the most effective inhibition
Discussion and Conclusion
DK, the PGD2 metabolite reverses the stimulation produced by several agonists, such as CPA, forskolin and carbachol. Calcium removal from the serosal solution has marked inhibitory effects on CPA and carbachol but not forskolin. Replacement of serosal calcium restores the responses. Pre-treatment with DK significantly inhibited responses to carbachol but not CPA or forskolin. This appeared to mimic the effects of verapamil. Nickel affected all 3 agonists. Replacement of serosal calcium in the absence of a stimulant had negligible effects. However prior treatment with CPA elicited clear concentration-responses to serosal additions of Ca2+. These effects were muted if CPA had been washed out prior to addition of Ca2+, suggesting the presence of a factor(s) that was easily removed. Under these conditions, DK markedly inhibited responses to added Ca2+, verapamil did not. Thus the prostanoid could affect either the production of this factor or more likely its effects on promoting serosal calcium entry. The effects of nickel were severe on all agonists and often appeared to be greater than the mere removal of serosal Ca2+, suggesting effects on non-specific cation channels. At least ONE locus for the inhibitory effects of PGD2 appears to be serosal Ca2+ entry.
References
- Rangachari, PK et al.(1992) Histamine potentiation by hydroxylamines: structure-activity relations; inhibition of diamine oxidase. Am. J. Physiol. (Gastrointest. Liver Physiol. 26), G632-641.
| Discussion Board | Previous Page | Your Poster Session |