Invited Symposium: Genital Sensation: CNS Targets and Functions in Females
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
INTRODUCTION
It has long been appreciated that genitosensory or vaginocervical (VCS) stimulation is required for the initiation of pregnancy in the rat, and that the necessary stimulation of the uterine cervix occurs when the estrous female receives intromissions from the male during the natural mating sequence.
In addition, VCS has been shown to induce changes in sexual receptivity which include an initial facilitation of lordosis intensity followed by a subsequent abbreviation of the duration of heat. Frequency and timing of the intromissions are important elements of the necessary mating stimulation, with both increasing numbers of intromissions and interintromission intervals contributing to the initiation of both these behavioral and the neuroendocrine changes.
Females mated under paced mating conditions in which they actively regulate the temporal characteristics of the intromissive stimulation require fewer numbers of intromissions to become pregnant/pseudopregnant than do females mated in standard lab tests in which the male, not the female, controls the timing of each intromission. Paced mating stimuli are also more effective than non-paced stimuli in abbreviating the duration of estrus, inducing LH and 3a-androstanediol secretion, and striatal DA release in females.
Therefore, for both behavioral and neuroendocrine processes, paced mating provides more effective VCS than does non-paced mating. Our recent approach to the study of how mating alters these behavioral and neuroendocrine processes is to compare FOS responses to paced and non-paced VCS to determine where and how VCS is processed in the CNS.
Surprising little information is available concerning the neural pathways involved in processing that afferent input received during mating which might selectively affect the induction of pseudopregnancy or alter estrus duration. It has been demonstrated that input from the pelvic nerve, the parasympathetic autonomic nerve which innervates the uterine cervix and vagina, bladder, rectum and colon, is required for both mating-induced pregnancy and estrus abbreviation, since transection of that nerve bilaterally completely eliminates the period of ovarian acyclicity which characterizes pregnancy or pseudopregnancy and prolongs the period of estrus. In addition, neonatal capsaicin treatment, which is known to result in degeneration of C-fibers, eliminates the ability of mating to induce pseudopregnancy. The sympathetic hypogastric nerve also innervates portions of the cervix as well as the caudal portions of the uterus, but it is not required for pseudopregnancy.
The experiments presented in the present paper were designed to determine whether selective responses to VCS occur within segments of the spinal cord which receive afferent input from the pelvic and hypogastric nerves and to determine, comparing paced and non-paced mating, whether some processing of VCS occurs at the spinal level which could contribute to the initiation of pseudopregnancy. Because we found that FOS responses were actually reduced in animals which received intromissions compared to those which received mounts-without-intromission, we carried out a further experiment to determine whether this reduction could have resulted from descending influences on the cord responsible for mating-induced analgesia.
EXPERIMENT 1: Effects of Mating on Spinal FOS
In the first experiment in this series, FOS immunocytochemistry was used to detect VCS- induced changes in the activity of spinal neurons at levels T11-S3. The pelvic nerve afferents are known to send afferents to segments L6-S2, while the hypogastric nerve inputs occur at T13-L1. Adult ovariectomized estrogen and progesterone-treated rats were sacrificed 1 hr after receiving mating stimulation from males which included 5 intromissions (5I) or 15 intromissions (15I), mounts-without-intromission by use of either vaginal masks (MO-M) or genitally-anesthetized (MO-L) males, or immediately after being removed from their home cages (HC). FOS counts were made using a camera lucida with the experimenter blind to the test treatment of the particular spinal cord being analyzed. The mean number of FOS-immunoreactive (FOS-IR) cells within the gray matter of the spinal cord at each segment are presented in Figure 1.
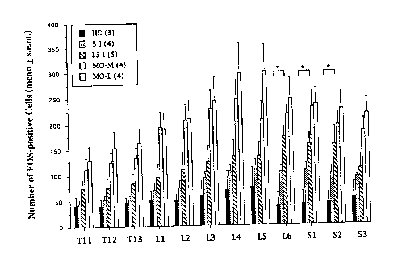 figure 1
figure 1
At all spinal levels, FOS labelling was lowest in the home cage group (50 � 22 cells), intermediate in the groups receiving intromissions (84 � 8 and 118 � 22 cells) and highest in groups receiving mounts-without-intromission stimulation (187 � 21 and 218 � 35 cells). Statistically significant changes among groups were present at all levels except T11. Significant increases above control levels following intromissive stimulation were observed at levels L6, S1 and S2. Surprisingly, both groups receiving mounts-without-intromission showed significantly higher numbers of FOS-positive cells than did one or both intromitted groups at all levels.
In order to more carefully examine whether the FOS responses occurred in second-order cells which would be expected because of their segmental location to receive input from the pelvic nerve, selected spinal segments were analyzed by Rexed's laminae at L6 (pelvic nerve) and L1 (hypogastric nerve). As seen in Figure 2, this analysis revealed that intromissive stimulation increased FOS labelling above control levels in laminae II, III, IV, V, and X at L6 (Fig. 2B) and in laminae I, II, V and X at S1. VCS did not increase labelling at L1 (Fig. 2A). The greater FOS responses seen in the mounts-without-intromission animals than in control or intromitted animals were apparent at L1, L6 and S1 within the same laminae (II, III, IV, V, and X).
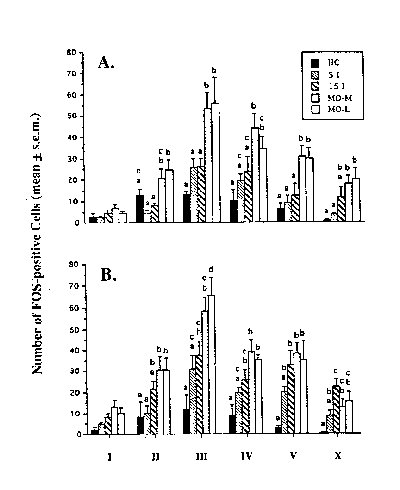 figure 2 Bars with different letters are significantly different from each other.
figure 2 Bars with different letters are significantly different from each other.
These results suggest 1) that stimulation of the uterine cervix initiates activity within L6-S2 neurons which receive pelvic nerve afferents and 2) that such stimulation suppresses activity at all levels within populations of neurons normally activated by cutaneous somatic inputs received from male mounts.
The suppression of FOS-IR below mounts-without-intromission levels by intromissions within laminae II, III, IV, V, and X at both L1 and L6 suggest a widespread intersegmental modulation of afferent input. The possible role of that modulation was explored in the subsequent experiments.
EXPERIMENT 2: Effects of Paced Mating on Spinal FOS
In the second experiment, we examined the effects of paced as opposed to non-paced mating on c-fos expression in the lumbar spinal cord. We compared the FOS response which occurred in ovariectomized hormone-primed females following 5 or 15 paced and non-paced intromissions. These numbers of intromissions were chosen because we had previously shown that 5 paced intromissions were significantly more likely to induce pseudopregnancy than was 5 non-paced intromissions, while 15 intromissions were shown to be equally effective when they were paced and when they were non-paced.
Thus, we expected that the FOS responses would differ between the two groups receiving 5 intromissions, and that the response in the 5 paced group would, based on the previous results, be significantly lower than that shown by the 5 non- paced group. The animals were mated or given mounts-without-intromission or control home cage treatment and were killed and their spinal cords taken 1 hr after mating onset. The results of this experiment are shown in Figure 3.
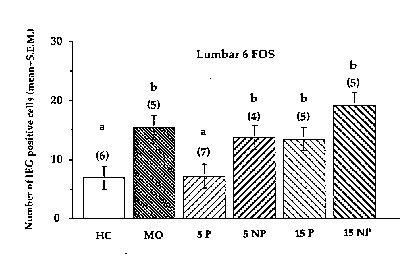 Figure 3. Bars with different letters are significantly different from each other.As can be seen, the
number of FOS-IR cells throughout the gray matter at L6 was significantly lower, as expected, in
the 5 paced group compared to the 3 other intromitted groups. As in the previous experiment,
the response among the 15 intromission groups were not different from the mounts-without-
intromission group.
Figure 3. Bars with different letters are significantly different from each other.As can be seen, the
number of FOS-IR cells throughout the gray matter at L6 was significantly lower, as expected, in
the 5 paced group compared to the 3 other intromitted groups. As in the previous experiment,
the response among the 15 intromission groups were not different from the mounts-without-
intromission group.
When the data were analyzed by lamina (Fig. 4), significant elevations due to mounts-without-intromission were again seen in laminae I & II, and significant increases in response to intromissions were seen in laminae VI (not analyzed in the previous study) and X. Although suppression due to intromissions was therefore observed, there was no particular site within the dorsal horn which showed reduced FOS responses which were selective to the 5 paced intromission group.
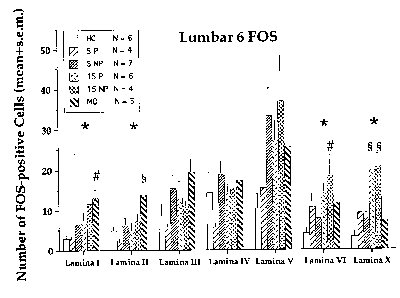 Figure 4. Asterisks indicate overall significant effect at this lamina. # - indicate increases above
the home cage group. $ - indicate significant increases above all other groups at that lamina.
Figure 4. Asterisks indicate overall significant effect at this lamina. # - indicate increases above
the home cage group. $ - indicate significant increases above all other groups at that lamina.
The results of this experiment demonstrate that paced mating is a more effective stimulus for suppression of FOS expression in L6 of the spinal cord than is non-paced mating. This increased effectiveness corresponds with the increased efficacy of paced mating on behavioral and neuroendocrine measures, and suggests that the intensity and timing of the paced mating stimulus may induce these effects by altering sensory processing at the initial point contact with the CNS. However, the lack of a specific laminar location for this suppression suggests that there may be a general dampening of responsiveness to the mating stimulation rather than a specific response which might be responsible for an analgesic response.
EXPERIMENT 3: Effects of Paced Mating on Tail Flick Latencies
As antinociceptive agents are known to suppress c-fos expression, VCS received during natural mating may be capable of initiating spinal and/or brain mechanisms of analgesia. In the last experiment, we examined whether paced as opposed to non- paced mating was more effective in inducing increases in tail-flick latency using the radiant heat method. Ovariectomized hormone-primed animals were given 4 baseline tests for tail-flick latency at 30 sec intervals immediately before mating, and 4 post-mating tail flick tests were performed immediately after completion of the mating sequence. The groups were: 5 intromissions during paced mating tests, 15 intromissions during paced mating tests, 5 intromissions during non-paced mating tests, and 15 intromissions during non-paced mating tests, and mounts-without- intromission animals. An additional group was given approximately the same number of intromissions as the 15 intromission groups, but post-mating tail flick tests were obtained immediately following the 2nd ejaculation received (2Ej group). The experiment was repeated during a second week, and the data from that week were used for analysis.
The post-mating tail flick tests were normalized as a percentage of the mean baseline tail flick latency and are presented in Figs. 5 & 6.
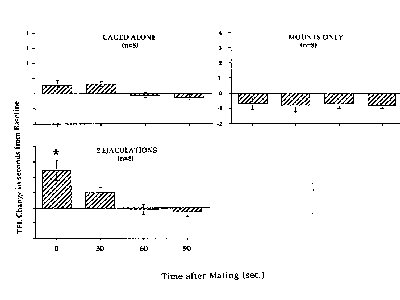 Figure 5. Asterisk indicates significant increase above baseline.
Figure 5. Asterisk indicates significant increase above baseline.
The results of this experiment were very interesting and in some ways unexpected. The tail-flick latency (TFL) for the 2Ej group was significantly greater than baseline, and the response,
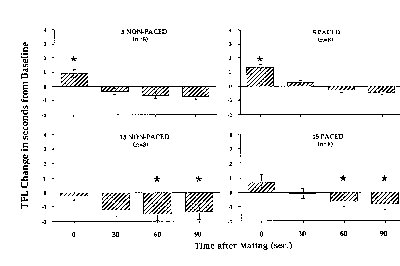 Figure 6. Asterisk indicates significant increase above baseline.
Figure 6. Asterisk indicates significant increase above baseline.
while brief, was strong. Similar increases in TFL were seen in both the paced and non-paced groups that received 5 intromissions, while no increase in TFL was observed in the groups receiving 15 intromissions or mounts-without-intromission. In the 15 intromission groups, significant hyperalgesia was observed during the latter tests. The duration of the mating tests did not differ significantly overall, although in general the 15 intromission tests were of longer duration than the 5 intromission tests. Therefore, these data show that the number of intromissions significantly affects the occurrence of VCS-induced analgesia, with fewer intromissions inducing analgesia and more intromissions inducing a hyperalgesia. There was no effect of paced vs. non-paced mating on this measure, so the overall changes in FOS-IR which were observed in the second experiment were not predictive of the analgesic response.
CONCLUSIONS
These studies demonstrate several interesting findings about spinal cord processing of VCS. First, intromissive stimuli received from males actually decreases the number of cells which label for FOS in the dorsal horn and lamina X below levels observed with mounts- without-intromission stimulation. This suppression is seen throughout the spinal gray and is observed when a low number of paced rather than non-paced intromissions are given. In contrast, analgesia, as measured by the tail flick method, is induced by low numbers of intromissions regardless of whether they are paced or non-paced. Hyperalgesia is observed following high numbers of intromissions, and, again, there is no effect of paced mating on this effect. These results suggest that the general suppression of FOS by intromissions is not associated with either an analgesic or hyperalgesic condition, but may represent segmental processing of reproductively- relevant afferent input from the pelvic nerve.
| Discussion Board | Previous Page | Your Symposium |