Invited Symposium: Genital Sensation: CNS Targets and Functions in Females
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Background
During the estrous cycle of female rats, the expression of sexual behavior is dependent upon an increase in estradiol levels followed by a surge of progesterone.
Sexual behavior is abolished by ovariectomy, and it is reinstated by the sequential treatment of estradiol followed one or more days later by progesterone.
Although estradiol treatment alone can induce sexual receptivity, when animals are treated subsequently with progesterone, lower doses of estradiol can be used, and the onset and termination of sexual behavior are more predictable. Progesterone also increases solicitation behavior in rats.
A great deal of work that has been done over the past twenty years supports the idea that intracellular progestin receptors (PRs), acting as transcription factors, are essential in mediating the effects of progesterone on sexual behaviors (2, 3).
While the appropriate underlying hormonal conditions are important for sexual receptivity, mating stimulation provided by the male also influences sexual behavior. Mating stimulation, including vaginocervical stimulation (VCS) causes many behavioral and endocrine changes in female rats, such as increases in lordosis, pseudopregnancy, and termination of sexual receptivity.
Females that are estradiol-primed, but not sexually receptive, show potentiation of sexual receptivity when repetitively mated with males. The mechanism by which this occurred was unknown; however it occurs in the absence of circulating progesterone, suggesting that progesterone is not involved.
Recent studies suggest that PRs may be activated in the absence of progesterone. Besides the in vitro evidence, intracerebroventricular infusion of dopaminergic agonists increases sexual receptivity in estradiol-primed rats, and treatment with progestin antagonists or antisense oligonucleotides directed at the progestin receptor mRNA blocks this facilitation (3). These studies suggest the exciting idea that steroid-independent activation of PRs may occur in vivo. We will discuss experiments which demonstrate that steroid-independent activation of PRs occurs after stimulation received during mating, and in particular in response to vaginocervical stimulation (VCS). Thus, genital stimulation may directly influence PR-dependent, genomic activity in the central nervous system.
PR-dependent changes in behavior
Repeated testing or exposure of estradiol-primed female rats to males increases sexual receptivity. As dopamine is released in the forebrain during mating (5), and as there is evidence that dopamine activates PRs (3), we tested the hypothesis that mating-induced potentiation of sexual receptivity in rats (that is, sexual behavior that occurs in the absence of circulating progesterone) requires PRs (2). Thus, increases in lordosis observed following repetitive mating or VCS in estradiol-primed female rats should be blocked by a progestin antagonist.
Ovariectomized/adrenalectomized female rats were primed with estradiol, and two days later were given manual VCS plus flank-perineum stimulation followed by mating with a sexually active male rat. One group of rats received the progestin antagonist, RU 486 prior to the stimulation, and one group received vehicle.
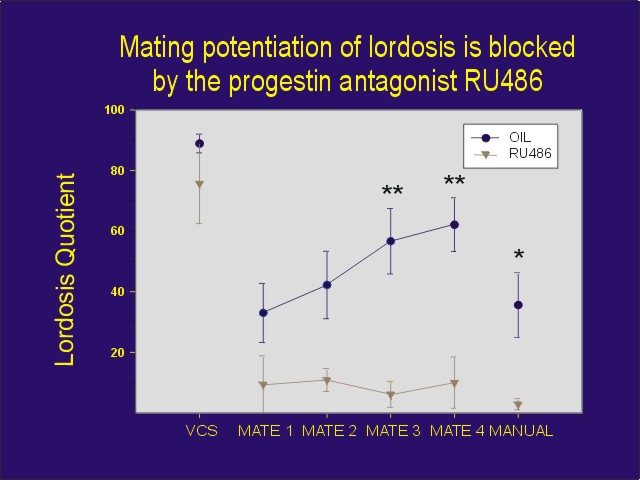
While manual VCS plus flank-perineum stimulation reliably induced lordosis at the time of stimulation, there was no difference in lordosis quotients between rats pretreated with the progestin antagonist RU 486 or oil vehicle. However, when the female rats were then placed in an arena with a sexually active male 15 min after VCS, RU 486-treated rats showed lower levels of sexual receptivity than the oil-treated controls (Figure 1). In subsequent mating tests, sexual receptivity increased by the third and fourth mating test in the oil-treated controls. In contrast, this increase was completely blocked by treatment with the progestin antagonist. Therefore, this experiment suggests that mating stimuli may activate PRs in a steroid-independent manner in the absence of progesterone resulting in changes in behavior.
 Click to enlarge
Figure 1: Effects of the progesterone antagonist RU 486 on sexual behavior of estradiol-primed female rats following VCS and repeated testing with a male. At the end of testing, rats were manually palpated by the experimenter.
Click to enlarge
Figure 1: Effects of the progesterone antagonist RU 486 on sexual behavior of estradiol-primed female rats following VCS and repeated testing with a male. At the end of testing, rats were manually palpated by the experimenter.
To determine if the behavioral consequences of blocking PRs were unique to the Type II progesterone antagonist RU 486, we repeated the experiment using the Type I progesterone antagonist, ZK 98299. As predicted, female rats receiving oil vehicle showed facilitation of sexual receptivity by the third and fourth mating test. However as with RU 486, the mating tests failed to facilitate sexual behavior in the rats treated with the progesterone antagonist.
Thus, mating stimulation facilitates sexual receptivity in estradiol-primed, ovariectomized/adrenalectomized female rats, and this facilitation is blocked by treatment with progesterone antagonists. This suggests that the increase in sexual receptivity that occurs in response to either somatosensory information provided by VCS or other stimuli associated with mating may be due to progesterone-independent activation of neuronal PRs.
It is unclear which sensory cues from the male are necessary to facilitate this effect. In a follow-up experiment, we asked the question of whether VCS was the relevant mating stimulus which caused potentiation of sexual receptivity.
VCS was prevented in one group by application of tape covering the vagina. While the unmasked controls showed a steady increase in lordosis over the course of the tests, masked group showed a much delayed an suppressed increase.
While these results suggest that VCS plays a critical role in the facilitation of lordosis, it is also likely that other sensory cues, such as olfactory stimuli, are involved as well.
Effects of mating stimulation on PRs
Besides facilitating the expression of sexual behavior under conditions of low levels of sexual behavior, mating also leads to the termination of sexual receptivity. We demonstrated earlier that progesterone appears to lead to termination of the period of sexual receptivity and to cause hyposensitivity to further facilitation by progesterone via down-regulation of neural PRs (2). Similarly, we hypothesized that mating too, might lead to down-regulation of PRs.
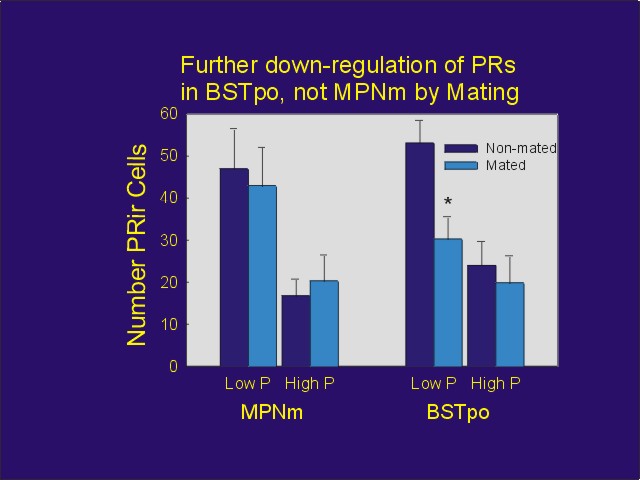
Ovariectomized/adrenalectomized female rats were injected with estradiol benzoate followed by either a low (0.5 mg) or high (5 mg) dose of progesterone. They were then administered manual VCS and mated with males. Twenty-four hours later, they were perfused , and their brains were immunostained for PRs using a PR antibody that is uninfluenced by hormonal occupation of the PR. Confirming earlier work, progesterone down-regulated PRs in all brain regions studied; furthermore, after the low dose of progesterone, mating caused additional down-regulation in some areas. For example, while PRs in the preoptic aspect of the bed nucleus of stria terminalis (BSTpo) were down-regulated by mating, those in the medial aspect of the medial preoptic nucleus (MPNm) were not (Figure 2).
 Click to enlarge
Figure 2: PR-IR counted unilaterally in either the medial aspect of the medial preoptic nucleus or the preoptic aspect of the bed nucleus of stria terminalis in ovariectomized rats treated with estradiol benzoate followed either by 0.5 mg progesterone (Low P) or 5 mg progesterone (high P). Mated animals were administered VCS and then mated with males 24 hours before perfusion.
Click to enlarge
Figure 2: PR-IR counted unilaterally in either the medial aspect of the medial preoptic nucleus or the preoptic aspect of the bed nucleus of stria terminalis in ovariectomized rats treated with estradiol benzoate followed either by 0.5 mg progesterone (Low P) or 5 mg progesterone (high P). Mated animals were administered VCS and then mated with males 24 hours before perfusion.
While the process by which mating down-regulates PRs is not known, it is possible that progesterone-independent activation of PRs, which accompanies mating, may then lead to subsequent down-regulation of PRs by a process similar to that by which progesterone does.
Because the data presented here suggest that genital stimulation results in increases in sexual receptivity by a ligand-independent process that involves PRs, we hypothesized that genital stimulation might induce Fos expression via activation of PRs.
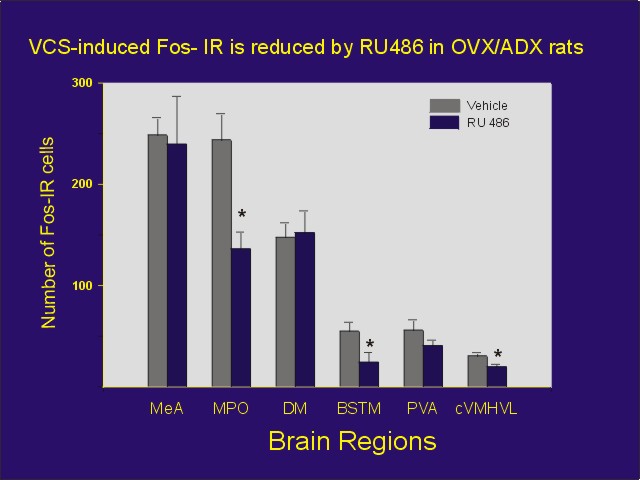
Estradiol-primed, ovariectomized/adrenalectomized female rats were administered VCS or control stimulation. One group of animals was administered the progestin antagonist, RU 486 prior to VCS. As predicted, RU 486, partially blocked the VCS-induced Fos expression in some brain areas, but not others (Figure 3).
 Click to enlarge
Figure 3: Number of Fos-IR cells per section for each area from estradiol-primed ovariectomized/adrenalectomized rats receiving manual VCS or control perineal stimulation. Rats received either 5 mg RU 486 or oil vehicle one hour prior to stimulation, and they were perfused one hour later. Abbreviations: MeA = medial amygdala, MPOA = medial preoptic area, DMH = dorsomedial hypothalamus, BSTM = bed nucleus of stria terminalis, PVA = anterior-periventricular thalamic nucleus, cVMHVL = caudal ventrolateral aspect of the ventromedial hypothalamus.
Click to enlarge
Figure 3: Number of Fos-IR cells per section for each area from estradiol-primed ovariectomized/adrenalectomized rats receiving manual VCS or control perineal stimulation. Rats received either 5 mg RU 486 or oil vehicle one hour prior to stimulation, and they were perfused one hour later. Abbreviations: MeA = medial amygdala, MPOA = medial preoptic area, DMH = dorsomedial hypothalamus, BSTM = bed nucleus of stria terminalis, PVA = anterior-periventricular thalamic nucleus, cVMHVL = caudal ventrolateral aspect of the ventromedial hypothalamus.
These results suggest that the VCS-induced Fos expression in some cells is dependent upon the availability of unoccupied PRs. They studies are consistent with the idea that some information provided by VCS or mating-stimulation activates neuronal PRs in the absence of progesterone.
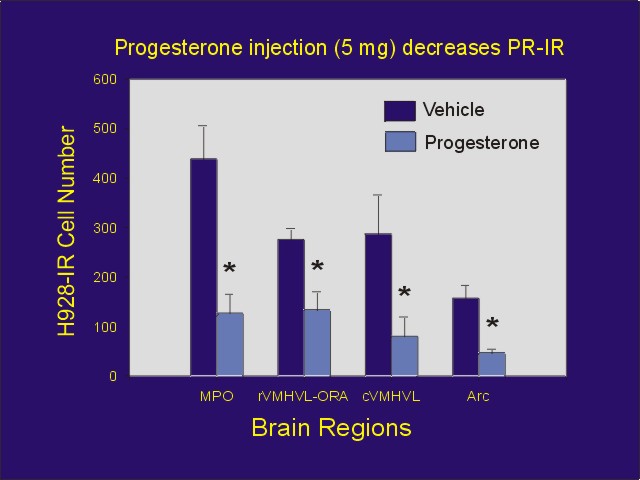
Although progesterone and afferent input conveying information from genital stimulation may each activate PRs, it is not known if they activate PRs by the same mechanism. In an attempt to determine if each type of activation has a similar effect on PRs, we either injected estradiol-primed ovariectomized/adrenalectomized rats with progesterone, or we administered VCS one hour prior to perfusion. We then immunostained brain sections using a PR antibody (H 928) that is influenced by occupation of the PR with ligand. Although progesterone decreased PR-immunostaining in all brain regions studied (Figure 4), VCS was without effect (Figure 5).
 Click to enlarge
Figure 4: Mean number of PR-IR cells (using H 928 antibody) after injection of estradiol benzoate followed by oil or progesterone. In addition to abbreviations defined in Figure 3: rVMHVL-ORA = ovarian steroid receptor-associated area associated with the rostral aspect of the ventrolateral ventromedial hypothalamus, Arc = arcuate nucleus
Click to enlarge
Figure 4: Mean number of PR-IR cells (using H 928 antibody) after injection of estradiol benzoate followed by oil or progesterone. In addition to abbreviations defined in Figure 3: rVMHVL-ORA = ovarian steroid receptor-associated area associated with the rostral aspect of the ventrolateral ventromedial hypothalamus, Arc = arcuate nucleus
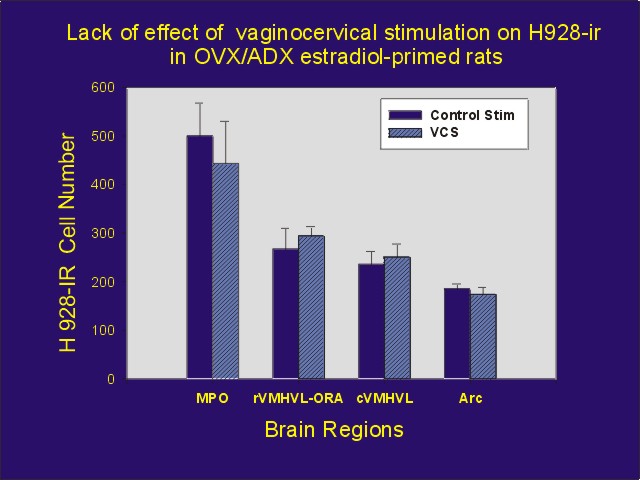 Click to enlarge
Figure 5: Mean number of PR-IR cells (using H 928 antibody) after injection of estradiol benzoate followed by VCS or control stimulation. In addition to abbreviations defined in Figure 3: rVMHVL-ORA = ovarian steroid receptor-associated area associated with the rostral aspect of the ventrolateral ventromedial hypothalamus, Arc = arcuate nucleus.
Click to enlarge
Figure 5: Mean number of PR-IR cells (using H 928 antibody) after injection of estradiol benzoate followed by VCS or control stimulation. In addition to abbreviations defined in Figure 3: rVMHVL-ORA = ovarian steroid receptor-associated area associated with the rostral aspect of the ventrolateral ventromedial hypothalamus, Arc = arcuate nucleus.
These results suggest that the process by which VCS activates PRs is different in some way from the process by which genital stimulation activates PRs. This raises the exciting possibility, that while hormones and neurotransmitters conveying information about genital stimulation may each be able to activate PRs, the PRs activated by the two routes may function in different ways. (Figure 5)
Discussion and Conclusions
The present results suggest that neuronal PRs may be activated by genital stimulation in a hormone-independent manner. These findings suggest the exciting idea of a cellular process by which genital stimulation influences steroid hormone receptor-dependent processes in the absence of circulating hormones. The resulting consequences of steroid receptor activation by these stimuli may produce changes in a wide variety of physiological, emotional, and behavioral events.
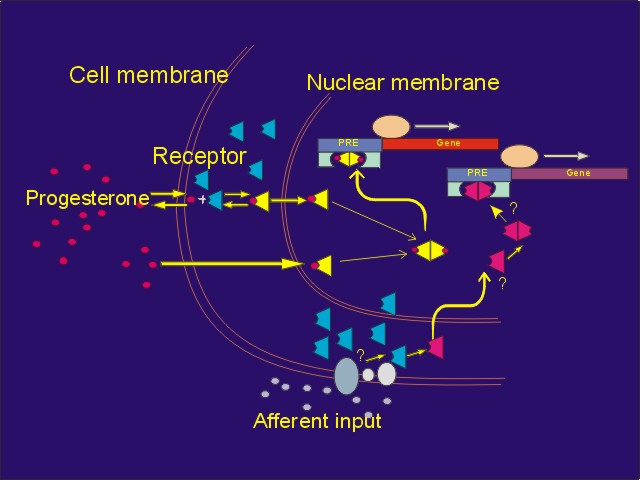 Click to enlarge
Figure 6: Convergence of afferent input and progesterone via PRs. Genital stimulation, acting through neurotransmitters, may activate PRs in some neurons, resulting in changes in the expression of progestin responsive genes. However, the process by which afferent input activates PRs differs from the process by which progesterone activates. PRs.
Click to enlarge
Figure 6: Convergence of afferent input and progesterone via PRs. Genital stimulation, acting through neurotransmitters, may activate PRs in some neurons, resulting in changes in the expression of progestin responsive genes. However, the process by which afferent input activates PRs differs from the process by which progesterone activates. PRs.
Mating stimulation, via PRs, can induce sexual behavior in animals that are not sexually receptive and who do not have circulating progesterone. However, under physiological conditions, it would be expected that progesterone acts on the receptors to cause the neurochemical changes that result in the onset of sexual receptivity. Mating stimulation also decreases the concentration of PRs in some neurons. Since we believe that the interactions of progesterone with PRs results in the down-regulation of PRs and termination of sexual receptivity, what function would mating-induced decreases in PRs have during physiological conditions in which rats are already exposed to progesterone? Perhaps genital and other social stimulation, acting through PRs, provides fine-tuning on the PR-dependent events, synchronizing the onset and termination of sexual behaviors to the most appropriate times. Furthermore, the effects of afferent input on PRs is localized to specific PR-containing neurons receiving specific inputs, while the effects of progesterone would not be expected to be specific to particular neurons.
We cannot yet define the neurotransmitters which cause activation of PRs. However, as mating stimuli increase dopamine release in the forebrain (5), dopamine agonists activate PRs (3), and a dopamine agonist induces Fos expression in some PR-containing neurons (4), it is possible that genital stimuli activate neuronal PRs via dopamine receptors. However, it is premature to rule out any other neurotransmitter or neuropeptide in this effect. As there is also a great deal of evidence for noradrenergic regulation of steroid hormone receptors (2), norepinephrine is also an excellent candidate for this regulation.
Some of the many questions raised by this work include the following:
If in fact mating stimulation can substitute for and/or potentiate the effects of progesterone on sexual behavior, then are there other physiological instances in which afferent input-induced activation of steroid hormone receptors influences behavior and physiology?
If genital stimulation can address PR-containing neurons in rats resulting in PR-dependent changes in behavior, consideration must be given to the possibility that such processes may occur in response to genital stimulation in all animals, including humans?
References
- Auger, AP, Moffatt, CA and Blaustein, JD (1997) Progesterone-independent activation of rat brain progestin receptors by reproductive stimuli. Endocrinology, 138: 511 - 514.
- Blaustein, JD and Olster, DH (1989) Gonadal steroid receptors and social behavior. In Advances in Comparative and Environmental Physiology, (J Balthazart, Ed.), pp 31 - 104, Springer Verlag, Berlin.
- Mani, SK, Blaustein, JD and O'Malley, BW. (1997) Progesterone receptor function from a behavioral perspective. Hormones and Behavior, 31: 244 - 255.
- Meredith, JM, Auger, AP and Blaustein, JD (1997) D1 dopamine receptor agonist (SKF-38393) induction of fos immunoreactivity in progestin receptor-containing areas of female rat brain. Journal of Neuroendocrinology, 9: 385 - 394.
- Vathy, I and Etgen, AM (1989) Hormonal activation of female sexual behavior is accompanied by hypothalamic norepinephrine release. Journal of Neuroendocrinology, 1: 383 - 388.
| Discussion Board | Previous Page | Your Symposium |