Invited Symposium: Oxidative Stress and the CNS
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
INTRODUCTION
Ascorbic acid, the ascorbate anion at physiological pH, is one of the most abundant low molecular weight antioxidants in the CNS (Lyrer et al. 1991). Ascorbate, unlike glutathione (GSH) (Orlowski & Karkowsky, 1976; Makar et al. 1994), is not synthesized in brain cells, but rather its cellular levels are maintained by active uptake. Whether ascorbate is synthesized in the liver, as in most mammals, or acquired from the diet, as in humans and a few other species (Chatterjee et al. 1975), it is carried to the brain via plasma. It is then actively transported across the blood-brain barrier (Spector and Lorenzo, 1973; Lam and Daniel, 1986) and subsequently taken up into brain cells from the extracellular fluid (Wilson, 1989; Kalir and Mytilineou, 1992; Rice et al. 1994). Each step in this transport process concentrates ascorbate: it is roughly 0.05 mM in plasma (Spector, 1977) and 0.5 mM in CSF (Stamford et al. 1984), with overall brain tissue levels of 2-3 mM (Milby et al. 1981; Rice et al. 1995).
In addition, we recently reported that ascorbate is differentially compartmentalized between neurons and glia, with an average intracellular concentration in neurons of 10 mM with slightly less than 1 mM in glia (Rice and Russo-Menna, 1998). Importantly, this difference is consistent with the difference in the estimated rate of oxidative metabolism in these cells, which is 10-fold higher in neurons than in glia (Siesjö 1980). This would mandate higher neuronal antioxidant levels to maintain antioxidant/oxidant balance and prevent oxidative stress. In addition, neurons and glia are affected differently when oxidative stress does occur, for example in cerebral ischemia or Parkinson's disease. Neurons are more susceptible than glia to ischemia/reperfusion injury (Kontos, 1989) and dopamine neurons degenerate while glia proliferate in Parkinson's (McGeer et al. 1989). GSH, on the other hand, is more concentrated in glia than in neurons (Slivka et al., 1987; Raps et al. 1989; Maker 1994; Rice and Russo-Menna, 1998).
In principle, ascorbate concentrations in neurons and glia could be determined using separate cell populations in culture. In practice, however, culture data have not provided consistent answers to the question of compartmentalization. In vitro, cellular levels of ascorbate, as well as GSH, are media-dependent, resulting in wide variation in reported contents (see Rice and Russo-Menna for further discussion and examples). At one extreme, when media are not supplemented with ascorbate or precursors of GSH, cells are antioxidant deficient. At the other extreme, levels much higher than physiological can be obtained. Such data suggest that quantitative estimates of cellular concentrations from cultured cells should be interpreted with caution.
We used an alternative approach to the question of ascorbate compartmentalization. We determined whole tissue levels of ascorbate (and GSH) in brain samples with varying, but known, neuron to glia ratios (Rice and Russo-Menna, 1998). First, we determined tissue contents during postnatal development in rat cortex and cerebellum, which have established patterns of neurogenesis and gliogenesis after birth. We then evaluated tissue contents in adult cortex from species with varying cortical neuron density. Cellular concentrations of ascorbate were calculated from these sets of quantitative data, in combination with appropriate intra- and extracellular volume fractions of the brain tissue under study. Ascorbate (and GSH) contents were determined using an HPLC method developed in this laboratory (Rice et al. 1995). Tissue was rapidly dissected, weighed, and frozen on dry ice within 10 min after decapitation. All samples were kept at –80 °C until analysis (Rice and Russo-Menna, 1998).
This paper summarizes and illustrates our evidence for differential compartmentalization of ascorbate between neurons and glia. Similar data for GSH distribution between these cells types is also included for comparison.
Regional and developmental data from rat
In adult rat CNS (Rice et al. 1995), ascorbate is more concentrated than GSH in all regions except optic nerve (Fig 1). The greatest contrast is between neuron-rich cerebral cortex and essentially neuron-free (Ransom et al. 1992) optic nerve. Whereas ascorbate content is roughly 4-fold higher cortex than optic nerve, GSH content differs by only 2-fold between gray and white matter. These data provided our first evidence that ascorbate might be predominantly localized in neurons, and were consistent with preferential localization of GSH in glia.
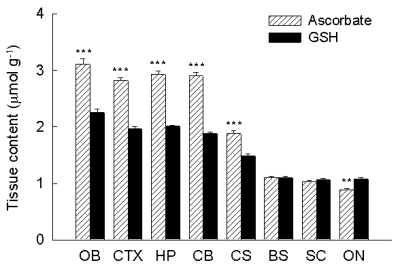 Fig. 1. Regional distribution of ascorbate and GSH in rat CNS.
Fig. 1. Regional distribution of ascorbate and GSH in rat CNS.
Data are mean ± SEM (significance of difference between ascorbate and GSH content is indicated by **p < 0.01 and ***p < 0.001). OB olfactory bulb, CTX cortex, HP hippocampus, CB cerebellum, SC superior colliculus, BS brainstem, SC spinal cord, ON optic nerve.
In the developing rat brain, regionally distinct changes in neuron:glia ratio occur during the first three postnatal weeks. In developing cerebral cortex, tissue ascorbate content is highest (Fig. 2A) and GSH lowest (Fig. 2B) shortly after birth, e.g. postnatal day 3 (P3), when cortical tissue is a nearly pure neuronal population with only few immature glial cells. Over the next few weeks, during the time of gliogenesis (Parnavelas et al, 1983) ascorbate content falls while GSH rises (Fig. 2).
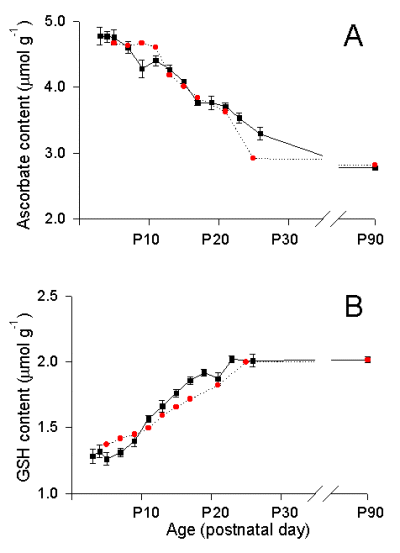 Fig. 2. Actual and calculated ascorbate (A) and GSH (B) contents in developing rat cortex.
Fig. 2. Actual and calculated ascorbate (A) and GSH (B) contents in developing rat cortex.
Tissue ascorbate and GSH contents at P3 were used to calculate ascorbate and GSH concentrations in neurons (see text and Rice and Russo-Menna, 1998). Black squares are experimental data; red circles are calculated ascorbate or GSH content. Experimental data are mean ± SEM; n = 8-14 samples per mean for P3-P26; n = 61 for P90.
In cerebellum, by contrast, ascorbate and GSH levels are constant from P3-P9 (Fig. 3). From P9, however, coincident with the onset of cerebellar granule cell proliferation (Altman, 1972), ascorbate levels increase markedly, reaching a maximum at P15-P17. Between P17 and P19, cerebellar ascorbate content falls by 10% , then continued to decrease at a similar rate over the next several days, until it reaches adult (e.g. P90) levels.
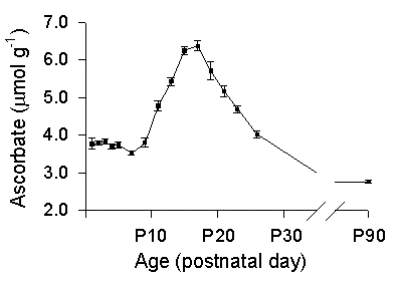 Fig. 3. Developmental changes in ascorbate content of in rat cerebellum.
Fig. 3. Developmental changes in ascorbate content of in rat cerebellum.
Ascorbate content begins to rise at P9, coincident with the onset of granule cell proliferation. Data from P15 were used to calculate neuronal ascorbate concentration (see text and Rice and Russo-Menna, 1998). Data are mean ± SEM; n = 6-15 samples per mean for P3-P26; n = 60 for P90.
The tissue ascorbate content of developing rat cortex at P3 and the ascorbate content of cerebellum at P15 can be assumed to represent levels in relatively pure neuronal populations. To calculate the intracellular concentration of ascorbate from these data requires that the fractional contributions of the extracellular compartment, the intracellular compartment, and the solid phase of the tissue be taken into account. Previously, Lehmenkuhler et al. (1993) determined that the extracellular volume fraction of rat cortex at P3 was 0.38. We determined that the solid phase fraction was 0.12 (Rice and Russo-Menna, 1998), such that the intracellular compartment would comprise 0.50 of the total tissue volume. Assuming an extracellular ascorbate concentration of 0.4 mM (supported by preliminary data), this permitted us to calculate an intracellular ascorbate of 9.2 mM. Intracellular ascorbate concentration in granule cell-rich cerebellum was similarly calculated to be 10.2 mM, based on an extracellular volume fraction of 0.25 (Rice et al. 1993) and solid phase fraction of 0.16, hence an intracellular volume fraction of 0.59. Using the tissue content of GSH on cortex at P3, we similarly calculated a neuronal GSH concentration of 2.5 mM. A detailed account of these calculations is given elsewhere (Rice and Russo-Menna, 1998). These data from neuron-rich populations in cortex and cerebellum not only demonstrate that ascorbate is preferentially localized in neurons, but indicate that it is highly concentrated at approximately 10 mM.
Ascorbate content and neuron density
If ascorbate were indeed highly concentrated in neurons and this occurred in adult as well as developing cells, one would expect a species dependence of adult cortical ascorbate content, with higher levels in species with greater neuron density (Tower and Elliott, 1952). Consistent with this hypothesis, cortical ascorbate content shows a strong, linear dependence on neuron density (Fig. 4; Rice and Russo-Menna, 1998). By contrast, GSH content is independent of neuron density (not illustrated). Ascorbate content exceeds that of GSH in mouse and rat brain, with a ratio of ascorbate to GSH of roughly 1.5 : 1 in these species. By contrast, GSH content is higher than ascorbate content in guinea pig, rabbit, and human cortical tissue (Russo-Menna, 1998), with ascorbate:GSH ratios of 0.4-0.7 : 1.
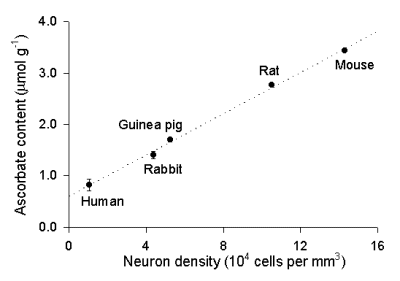 Fig. 4. Dependence of ascorbate content in cerebral cortex on cortical neuron density.
Fig. 4. Dependence of ascorbate content in cerebral cortex on cortical neuron density.
Ascorbate content increases linearly (r 2 = 0.997) with increasing neuron density across species. Data are mean SEM; n = 9-61 samples per mean. The y-intercept was used to estimate ascorbate concentration in glia (see text and Rice and Russo-Menna, 1998; neuron density data are from Tower and Elliott, 1952).
The y-intercept of the plot of ascorbate content versus neuron density (Fig. 4) indicates tissue ascorbate levels when neuron density is zero, i.e. a pure glial population. Again, with information about the intra- and extracellular volume fractions and the solid fraction, intracellular concentration, can be estimated. The ascorbate concentration in glia, therefore can be calculated to be 0.9 mM, using a extracellular volume fraction of 0.195 (Cserr et al., 1991; Lehmenkuhler et al, 1993), a solid phase of 0.198 and an intracellular fraction of 0.61 (Rice and Russo-Menna, 1998). From a similar plot of GSH vs. neuron density, we calculated a glial GSH concentration of 3.8 mM.
Summary and significance
In a test of the reliability of these data from ascorbate concentrations in neurons (10 mM) and in glia (0.9 mM), we used the data to predict the pattern of changes in tissue ascorbate content in developing rat cortex. Illustrated in Fig. 2 are calculated levels of ascorbate (Fig. 2A; red circles), along with experimental data from developing cortex (closed squares). In this model, we estimated developmental changes in intra- and extracellular volume fractions, as well as known values of neuron to glia ratio during development (Parnavelas et al. 1983). Full details of the calculations and certain assumptions are given in Rice and Russo-Menna (1998). Similarly, using our estimates of GSH concentrations in neurons (2.5 mM) and glia (3.8 mM), with the same calculated changes intra-glial and intra-neuronal volume fractions during development, we were also able to model the pattern of GSH changes in developing cortex to adulthood (Fig. 2B; red circles).
As noted earlier, the 10-fold difference in ascorbate content between neurons and glia approximates the difference in the rates of oxidative metabolism between these cell types, consistent with an important role for ascorbate in protection against oxidative stress. Indeed, ascorbate neuroprotection has been demonstrated in several recent studies, both in vitro and in vivo. Furthermore, ascorbate levels, but not those of GSH, are elevated in the CNS of anoxia-tolerant pond turtles (Rice et al. 1995), animals that routinely undergo hypoxia/reoxygenation during diving behavior. Although ascorbate can act as a pro-oxidant in vitro, as recently reviewed by Halliwell (1996), the present findings, taken with other data, argue strongly against normally pro-oxidant actions in vivo.
References
- Altman J. (1972b) Postnatal development of the cerebellar cortex of the rat: III. Maturation of the components of the granular layer. J. Comp. Neurol. 145, 465-514.
- Chatterjee I. B., Majumbar A. K., Nandi B. K. and Subramanian N. (1975) Synthesis and some major functions of vitamin C in animals. Ann. N.Y. Acad. Sci. 258, 24-47.
- Cserr H. F., DePasquale M., Nicholson C., Patlak C., Pettigrew K., and Rice M. E. (1991) Extracellular volume decreases while cell volume is maintained by uptake of ions in rat cerebral cortex during acute hypernatremia. J. Physiol. 442, 277-295.
- Halliwell B. (1996) Vitamin C: antioxidant or pro-oxidant in vivo. Free Rad. Res. 25, 439-454.
- Kalir H. H. and Mytilenou C. (1992) Ascorbic acid in mesencephalic cultures: effects on dopaminergic neuron development. J. Neurochem. 57, 458-464.
- Kontos H. A. (1989) Oxygen radicals in CNS damage. Chem.-Biol. Interactions. 72, 229-255.
- Lam D. K. C. and Daniel P. M. (1986) The influx of ascorbic acid into the rat's brain. Quart. J. Exp. Physiol. 71, 483-489.
- Lehmenkuhler A., Sykova E., Svoboda J., Zilles K. and Nicholson C. (1993) extracellular space parameters in the rat neocortex and subcortical white matter during postnatal development determined by diffusion analysis. Neuroscience 55, 339-351.
- Lyrer P., Landolt H., Kabiersch A., Langemann H. and Kaeser H. (1991) Levels of low molecular weight scavengers in the rat brain during focal ischemia. Brain Res. 567, 317-320.
- Makar T. K., Nedergaard M., Preuss A., Gebard A. S., Perumal A. S. and Cooper A. J. L. (1994) Vitamin E, ascorbate, glutathione, glutathione disulfide, and enzymes of oxidative metabolism in cultures of chick astrocytes and neurons: evidence that astrocytes play an important role in oxidative processes in the brain. J. Neurochem. 62, 45-53.
- McGeer P. L., Itagaki S., Akiyama H., McGeer E. G. (1989) Comparison of neuronal loss in Parkinson's disease and aging. Adv. Neurol. 36, 25-34.
- Milby K., Mefford I. N., Chey W. and Adams R. N. (1981) In vitro and in vivo depolarization coupled efflux of ascorbic acid in rat brain preparations. Brain Res. Bull. 7, 237-242.
- Orlowski M. and Karkowsky A. (1976) Glutathione metabolism and some possible functions of glutathione in the nervous system. Int. Rev. Neurobiol. 19, 75-121.
- Parnavelas J. G., Luder R., Pollard S. G., Sullivan K. and Lieberman A. R. (1983) A quantitative and qualitative ultrastructural study of glial cells in the developing visual cortex of the rat. Phil. Trans. R. Soc. (Lond.) 301, 55-84.
- Ransom B. R. Walz W., Davis P. K. and Carlini W. G. (1992) Anoxia-induced changes in extracellular K+ and pH in mammalian central white matter. J. Cereb. Blood Flow. Metab. 12, 593-602.
- Raps S. P., Lai J. C. K., Hertz L., and Cooper A. J. L. (1989) Glutathione is present in high concentrations in cultured astrocytes but not in cultured neurons. Brain Res. 493, 398-401.
- Rice M. E., Lee E. J. K. and Choy Y. (1995) High levels of ascorbic acid, not glutathione, in the CNS of anoxia-tolerant reptiles, in contrast to levels in anoxia-intolerant species. J. Neurochem. 64, 1790-1799.
- Rice M. E., Perez-Pinzon M. A. and Lee E. J. K. (1994) Ascorbic acid, but not glutathione, is taken up by brain slices and preserves cell morphology. J. Neurophysiol. 71, 1591-1596.
- Rice, M. E. and Russo-Menna, I. (1998) Differential compartmentalization of brain ascorbate and glutathione between neurons and glia. Neuroscience 82, 1213-1223.
- Siesjo B. K. (1980) Brain Energy Metabolism. pp. 131-150. Wiley, New York.
- Slivka A., Mytilineou C. and Cohen G. (1987) Histochemical evaluation of glutathione in brain. Brain Res. 409, 275-84.
- Spector, R. (1977) Vitamin homeostasis in the central nervous system. N. Engl. J. Med. 296: 1293-1398.
- Spector R. and Lorenzo A. V. (1973) Ascorbic acid homeostasis in the central nervous system. Am. J. Physiol. 225, 757-763.
- Stamford J. A., Kruk Z. L. and Millar J. (1984) Regional differences in extracellular ascorbic acid levels in the rat brain determined by high speed cyclic voltammetry. Brain Res. 299, 289-295
- Tower D. B. and Elliott K. A. C. (1952) Activity of the acetylcholine system in cerebral cortex of various unanesthetized animals. Am J. Physiol. 168, 747-759.
- Wilson J. X. (1989) Ascorbic acid uptake by a high-affinity sodium-dependent mechanism in cultured rat astrocytes. J. Neurochem. 53, 1064-1071.
| Discussion Board | Previous Page | Your Symposium |