Cardiovascular Diseases Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
An alpha1B-knockout (KO) mouse has recently been shown to have markedly reduced pressor responses to phenylephrine and reduced adrenergic receptor (AR) number in the liver, heart and brain (Cavalli et.al., 1997). alpha1B-AR's have also been suggested to be involved in the growth (hypertrophy) of smooth muscle cells (Chen et al., 1995) and are involved in the contraction of cutaneous arteries (Smith et.al., 1997) and spleen (Burt et al., 1995; Eltze). Mallard et al., (1992) have suggested that alpha1B-AR's may be confined to the synaptic region whereas Stassen et.al., (1998) have shown a positive relationship between degree of sympathetic innervation and density of alpha1A-AR's. To determine the importance of the alpha1B-AR in sympathetic transmission, we have examined the response to neuronally released noradrenaline (NA) on segments of isolated tail artery from wild type (WT) and KO mice. In addition we have used confocal microscopy to look for structural differences in WT and KO arterial segments
Materials and Methods
Male C57-Black WT and KO mice (35-40g) were killed by stunning followed by exsanguination. Arterial segments (2-3mm) were removed from the middle of the tail, mounted and normalised (0.9L100) on a wire myograph. Cumulative concentration response curves (CRC; 1nM-10µM) or frequency response curves (4, 8, 16, 32 & 64Hz; 0.1ms PW; 35v; 1 sec. train) were constructed in the presence and absence of chloroethylclonidine (CEC; 10µM for 30 mins followed by washing). An additional low frequency long duration stimulus was tested (4Hz for 10 seconds.)
Total RNA was extracted from WT and KO mouse brain according to the acid phenol/guanidinium thiocyanate method of Chomczynski & Sacchi (1987). Samples of 10µg of RNA were reverse transcribed using a first strand cDNA synthesis kit (Pharmacia). PCR reactions using Taq polimerase were carried out using the following primers: alpha1B sense, CCACTCTAAGAACTTTCATGAGGACACC; alpha1B antisense, ATGCAGCTGCCACTGTCATCCAGAGAGT. Amplifications were performed in a progene thermocycler. Cycles were as follows: 94oC for 5 mins. (1 cycle); 94oC for 2 mins., 56oC for 1 min., 72oC for 2 mins.(40 cylces); 72oC for 10 mins. (1 cycle). In addition, alpha1B-AR cDNA (200ng) was also subjected to PCR. Reaction products were separated by 1% agarose gel electrophoresis.
A Noran Laser Scanning Confocal Microscope (LSCM) was used to collect images of adventitial cell nuclei stained with Hoescht 33342 (10ug/ml; 364 ex./400 em; x25).
Results
RTPCR confirmed the absence of the alpha1B-message from brain tissue in the KO mice
 Figure 1. PCR products (10µl) from wild type (lanes 1,3) and KO (lanes 2,4), along with cDNA encoding mouse a1B-AR (lane 5) were subjected to agarose gel electrophoresis as described in methods. PCR amplified product of 410bp was detected by ethidium bromide fluorescence on a 300nm UV transilluminator.
Figure 1. PCR products (10µl) from wild type (lanes 1,3) and KO (lanes 2,4), along with cDNA encoding mouse a1B-AR (lane 5) were subjected to agarose gel electrophoresis as described in methods. PCR amplified product of 410bp was detected by ethidium bromide fluorescence on a 300nm UV transilluminator.
In WT mice the CRC to NA was shifted to the right by the high affinity alpha1-AR antagonist YM12617 (pKB 9.41) but not by the high affinity alpha2-AR antagonist delequamine (pKB 6.02).
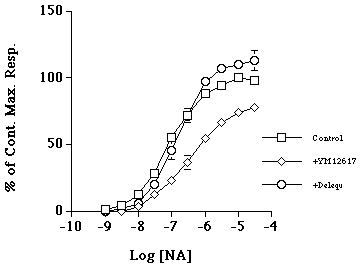 Figure 2. Concentration response curves to noradrenaline (square) in WT mouse tail artery in the presence of the high affinity a1-AR antagonist YM12617 (tamsulosin; diamond) and the alpha2-AR antagonist delequamine (circle). Data represent mean of n=4 ± s.e.m.
Figure 2. Concentration response curves to noradrenaline (square) in WT mouse tail artery in the presence of the high affinity a1-AR antagonist YM12617 (tamsulosin; diamond) and the alpha2-AR antagonist delequamine (circle). Data represent mean of n=4 ± s.e.m.
The EC50 values for NA in WT (6.93) and KO (6.64) were not significantly different (Fig 3a). CEC caused a 20% depression in the maximum response to NA in both WT (EC50 6.27) and KO (EC50 6.46) (Fig 3b&c).
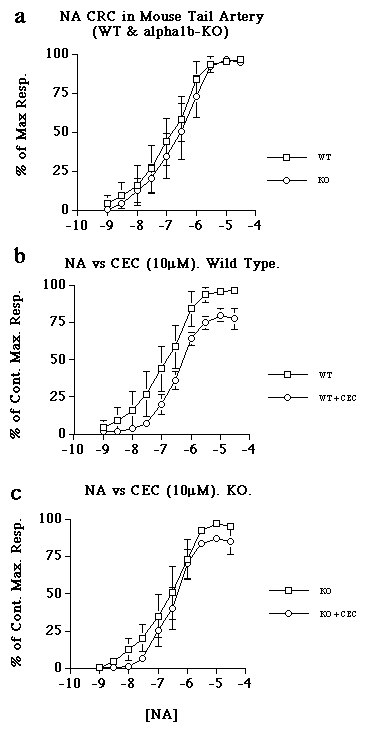 Figure 3. Concentration response curves to noradrenaline in WT and KO mouse tail artery in the presence and absence of chloroethylclonidine (CEC). a) Comparison of control curves for NA in WT and KO. b) the effect of CEC (10µM) versus NA in WT tail artery. c) the effect of CEC (10µM) versus NA in KO tail artery. Data represent mean of n=4 ± s.e.m.
Figure 3. Concentration response curves to noradrenaline in WT and KO mouse tail artery in the presence and absence of chloroethylclonidine (CEC). a) Comparison of control curves for NA in WT and KO. b) the effect of CEC (10µM) versus NA in WT tail artery. c) the effect of CEC (10µM) versus NA in KO tail artery. Data represent mean of n=4 ± s.e.m.
Responses to electrical field stimulation (EFS) were transient, normally returning to baseline within 20 seconds (Fig. 4a&b).
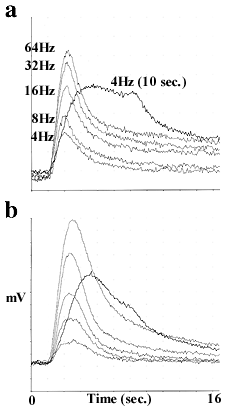 Figure 4. Representative tracings of responses to electrical field stimulation in the tail arteries taken from a) WT and b) KO mice. Stimulus parameters (delivered at the 2 seconds time point) are given in methods.
Figure 4. Representative tracings of responses to electrical field stimulation in the tail arteries taken from a) WT and b) KO mice. Stimulus parameters (delivered at the 2 seconds time point) are given in methods.
The responses to low frequency stimulation were significantly smaller in the KO (4-16Hz, p<0.1) (Fig 5a). Time to EFS induced maximum response ranged from 2.5 - 2.8sec. WT and 2.9 - 3.6sec in KO (p<0.1 for 4&8Hz) (Fig 5b). A 4Hz (10sec train) reached maximum amplitude at 6.9 seconds and 9.15 seconds for WT and KO respectively (p<0.1) (Fig 5b). The slope of the KO response (mg/sec) was reduced at low frequencies (4-16Hz; p<0.1) (Fig 5c).
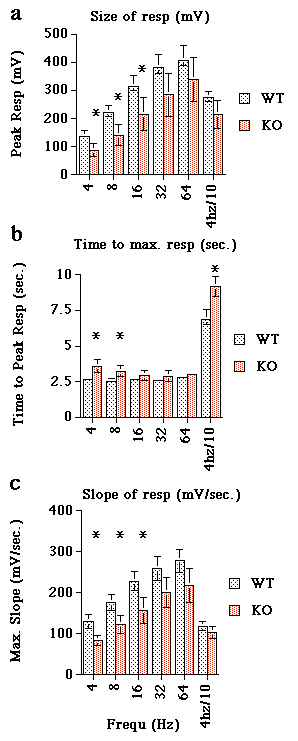 Figure 5. Analysis of 4 experiments of the type shown in figure 4. a) Comparison of the magnitude of responses of WT (light hatch) and KO (dark hatch) arteries to increasing frequencies of electrical field stimulation. b) The data shows the average time taken to reach peak amplitude (i.e. maximum response). c) Average slope expressed as mV per second for each frequency tested. Data represent mean of n=4 ± s.e.m. *=p<0.1
Figure 5. Analysis of 4 experiments of the type shown in figure 4. a) Comparison of the magnitude of responses of WT (light hatch) and KO (dark hatch) arteries to increasing frequencies of electrical field stimulation. b) The data shows the average time taken to reach peak amplitude (i.e. maximum response). c) Average slope expressed as mV per second for each frequency tested. Data represent mean of n=4 ± s.e.m. *=p<0.1
CEC inhibited the responses to EFS in both WT and KO arteries (Fig 6a&b).
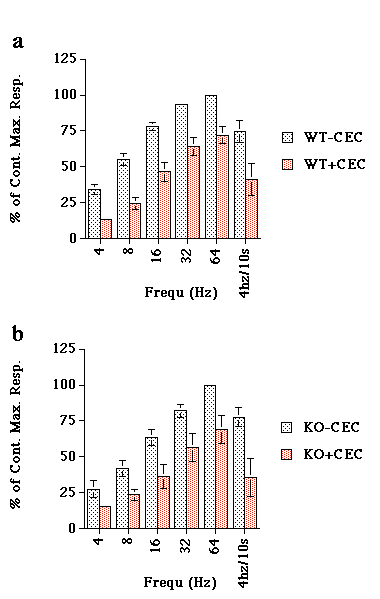 Figure 6. The effect of chloroethylclonidine (CEC; 10µM) versus electrical field stimulation of a) WT and b) KO mouse tail artery. Data are expressed as percent of maximum response achieved. Data represent mean of n=4.
Figure 6. The effect of chloroethylclonidine (CEC; 10µM) versus electrical field stimulation of a) WT and b) KO mouse tail artery. Data are expressed as percent of maximum response achieved. Data represent mean of n=4.
LSCM analysis of tail artery adventitia revealed a lower cell density in the KO compared with WT (Fig 7a&b)
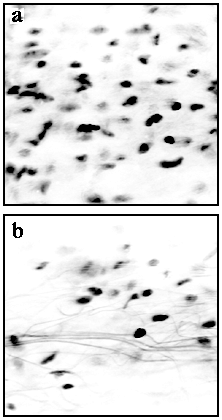 Figure 7. Representative confocal images of the cell nuclei (black dots) in the adventitia of a) WT and b) KO mouse tail artery. Arteries were stained with a nuclear stain and imaged using a confocal microscope (see methods). Three sets of arteries from different animals revealed identical results (i.e. reduced cell number and increased amount of connective tissue).
Figure 7. Representative confocal images of the cell nuclei (black dots) in the adventitia of a) WT and b) KO mouse tail artery. Arteries were stained with a nuclear stain and imaged using a confocal microscope (see methods). Three sets of arteries from different animals revealed identical results (i.e. reduced cell number and increased amount of connective tissue).
Discussion and Conclusion
These results suggest that the alpha1B-AR may play a role in sympathetic neurotransmission. Furthermore, the reduced cell number in the adventitia of the KO arteries supports the suggestion by Chen et al., that the alpha1B-AR may be involved in regulating growth. The use of single (and multiple) knockout mouse models provides a means of studying adrenergic receptor subtype function where the complicating factors of receptor subtype interactions can be eliminated or reduced.
References
Burt, R.P., et al., (1995). Br. J. Pharmacol. , 115; 467-475.
Cavalli, A. et al., (1997). Proc. Natl. Acad. Sci. USA, 94; 11589-11594
Chen, L. et al., (1995). J. Biol. Chem., 270 No. 52; 1-9
Chomczynski, P. & Sachi, N. (1987). Anal Biochem ., 162; 156-159.
Eltze, M. (1996). Eur. J. Pharmacol., 311; 187-198.
Mallard, N.J., et al., (1992). Br. J. Pharmacol., 105; 727-731.
Smith, K.M., et al., (1997). Br. J. Pharmacol. , 122; 825-832.
| Discussion Board | Previous Page | Your Poster Session |