Immunology & Immunological Disorders Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Results
EFFECT OF FAT CLEARANCE ON THE SEQUESTRATION OF SUSPENDED BACTERIA BY BREAST-MILK SAMPLES
Fat clearance of breast-milk by centrifugation reduced the bacteria count recovered from suspension to the levels comparable to that of control saline (Fig. 1).
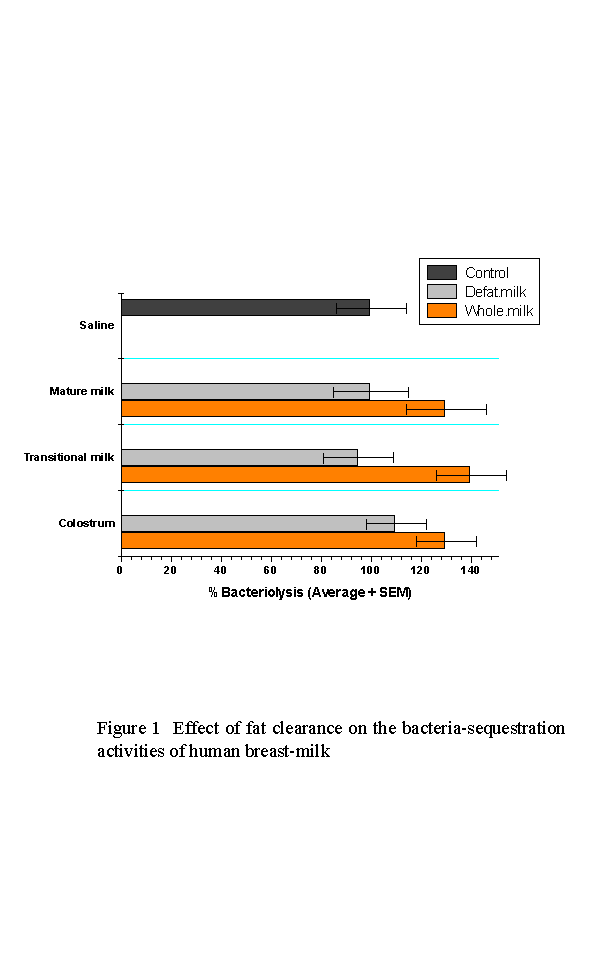
This difference was taken as a measure of sequestration by MFGM. The effects of fat clearance is on the average greater for both the transitional and mature milk samples than for colostrum.
EFFECT OF FREEZE-THAWING CYCLES ON THE SEQUESTRATION OF SUSPENDED BACTERIA BY HUMAN BREAST-MILK
Freeze-thawing of whole milk by repeated freezing at -20°C and subsequent thawing are known to be destructive to biological membranes (10). Frozen whole-milk at -20°C showed a fairly constant level of bacterial sequestrating capacities, slightly lower than those of fresh milk and those stored at -70°C, irrespective of repeated freeze-thawing (Fig 2).
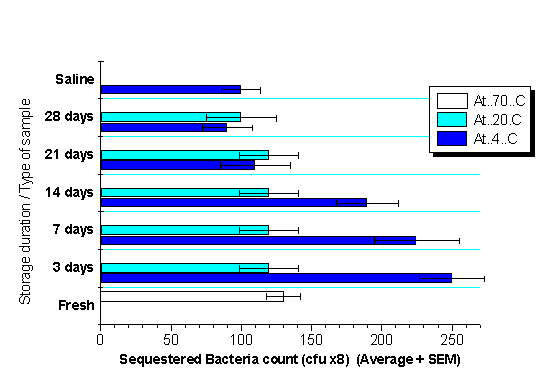 Figure 2 Comparison of the effects of refrigeration on the bactericidal and sequestration activities of human colostrum.
Figure 2 Comparison of the effects of refrigeration on the bactericidal and sequestration activities of human colostrum.
This failure of enhanced sequestration at -20°C might suggest that freezing either alters the surface of the MFGM, so that they are unable to bind to the limited level of Cp activation products in the milk samples, or the freeze-thawing procedures rapidly lead to their destruction.
This observation provides an evidence for the contribution of intact MFGM in binding and sequestration of suspended bacteria by HBM.
INCREASED SEQUESTRATION OF SUSPENDED BACTERIA BY HUMAN BREAST-MILK IN THE PRESENCE OF INCREASED ACTIVATED COMPLEMENT PRODUCTS
The sequestration abilities of whole-milk samples stored by refrigeration at 4°C in the absence of anticoagulants, under in-vitro conditions which favoured Cp activation, was compared to those frozen at -20°C or -70°C. It was observed that the refrigerated samples sequestered significantly more viable bacteria within the first two weeks of storage than fresh milk samples; followed by a progressive fall, gradually approaching the level cultured from the normal saline control (Fig 2).
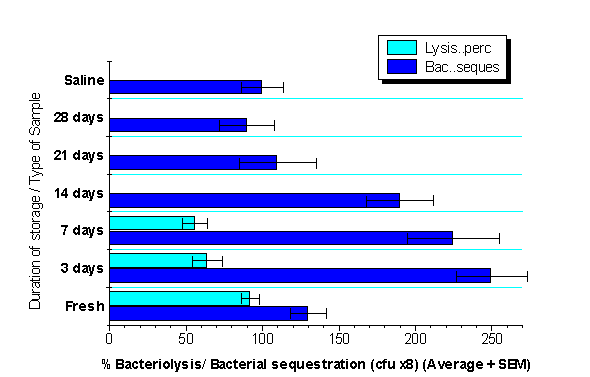 Figure 3 Comparison of the effects of refrigeration on the bactericidal and sequestration activities of human colostrum.
Figure 3 Comparison of the effects of refrigeration on the bactericidal and sequestration activities of human colostrum.
This enhancement of bacteria sequestration could be attributed to the presence of accumulation of activated Cp fragments in the stored milk, since they were not obtained in similarly stored milk samples containing EDTA. The other possible explanation for this observation might be the effect of FFA released by ongoing lipolysis, which is also inhibited by EDTA (6). This however seems unlikely since milk samples frozen at -20°C, containing higher levels of FFA compared to those stored at -70°C, did not demonstrate a corresponding enhancement of bacteria sequestration.
Previous studies have confirmed the accumulation of Cp split products, resulting from ongoing Cp activation, in biological samples stored at temperatures below -70°C (27). The accumulation of these split products in HBM stored by either refrigeration or freezing in the absence of EDTA have also been documented, using the same ELISA technique used for the deposition of C3 fragments (results not shown). The rapidly declining degree of bacteriolysis during the period of refrigeration, within the first 48 hours in mature milk and within one week in colostrum, further attest to ongoing losses of native Cp components through non-specific activation, and simultaneous accumulation of activated fragments (Fig. 3).
Figure 4 Comparison of the effects of refrigeration on the bactericidal and sequestration activities of human colostrum.
The subsequent fall in the sequestrating abilities of refrigerated milk samples might reflect the presence of ongoing in-vitro lipolysis associated with ultimate loss of intact MFGM (6), as well as the ultimate degradation of the Cp-activation products. This further confirms the necessity of an intact MFGM, independent of the presence of activated Cp fragments, to effect bacterial sequestration by milk samples.
ACTIVATION AND DEPOSITION OF ACTIVATED COMPLEMENT C3 FRAGMENT ON KILLED ESCHERICHIA COLI
The levels of complement derived opsonins-activated C3 (C3b, iC3b, C3dg), deposited on a solid-phase killed bacteria was estimated, using ELISA techniques, with help of an antibody (13/15) capable of recognizing these activated products of Cp activation (30). This provided evidence for the in-vitro capacity of activating the HBM Cp system.
The maximal levels of C3 opsonins deposition from milk samples were obtained by 1:2 dilutions of whole-milk in detergent-containing buffer (PBS-Tw), in comparison to dilutions in only PBS, SGVB++ or Mg++EGTA buffers (Table 1). The detergent-buffer probably displaces membrane-bound fragments by disrupting the MFGM through dissolution of the fat content. This suggests that the MFGM strongly competes with the solid-phase bacteria for the deposition of opsonins. Fat clearance of the milk samples however failed to restore the deposition of opsonins to levels comparable to that of whole-milk in detergent buffer, suggesting that a significant portion of the native Cp components might also have been lost through centrifugation.
The opsonins deposition found in the milk that had been heated for 30 minutes at 56°C probably demonstrates the presence of C3 fragments pre-formed in-vivo, an indication of possible physiological function in the lactating mammary gland. Table 1 Deposition of activated C3§ (opsonins) on killed E. coli 0111 bacteria by human breast-milk a c (results in µg/ml)
COLOSTRUM TRANSITIONAL MILK
-----------------------------------------------------------------------
Post-partum duration of lactation 1-4 days 5-30 days
Heatedb wholemilk 65 ± 10* 70 ± 5.5*
Wholemilk in PBS 75 ± 28 87 ± 12.8
Wholemilk in PBS-Tw buffer 167 ± 34 110 ± 25
Wholemilk in GVB2+ buffer 3 ± 2.2 --------
Partially de-fatted milk
(x500-3000g) in dGVB2+ buffer 84 ± 12 42 ± 6
Wholemilk in Mg-EGTA buffer ------- 50 ± 6.8
Partially de-fatted milk
(x500-3000g) in eMg-EGTA buffer 14 ± 5.6 64 ± 8.4
§ including C3b, iC3b, C3g and C3dg* Real value of deposited opsonin with no subtraction of the background opsonization.
a All values obtained after subtraction of the background level of opsonization (except those marked by * symbol)
b Whole-milk heated to 56°C for 30 minutes
c Average of two readings from each of two samples
d Supports the classical pathway of C activation
e Supports the alternative pathway of C activation
| <= Materials & Methods | RESULTS | Discussion & Conclussions => |
| Discussion Board | Next Page | Your Poster Session |