| INABIS '98 Home Page | Your Poster Session | Related Symposia & Posters | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Effects
OF Drought (Water Stress) On Growth And Photosynthetic Capacity Of Cotton
(Gossypium hirsutum L.)
Introduction
Drought is
a serious problem that affects many regions of the world, decreasing the
photosynthetic rate of crops and limiting the productivity world-wide.
Therefore, water availability is an essential factor influencing agriculture.
Growth and
photosynthesis are two of the most important processes abolished, partially
or completely, by water stress (Kramer and Boyer,1995), and both of them
are major cause of decreased crop yield.
Cotton crops
need irrigated lands to reach high productivities. Therefore, it is very
important to know how much and how often cotton plants should be irrigated
to complete a successful development.
The aim of
this work was to study how cotton plants are affected by different levels
of water stress. We particularly investigated when the first signs of stress
appear, which are the most sensitive developmental stages (seeds, seedlings
or young plants), and what organs (cotyledons or leaves) are the most affected
by drought.
Materials & Methods
Gossypium hirsutum L.cv. palma was grown in a controlled environment
chamber at day/night temperature 28/21°C, relative humidity 70/80%
and with a photoperiod of 16 h per day. The photosynthetic photon flux
was 200 micromol m-2 s-1 at plant height.
Plants were
grown hydroponically in plastic pots containing a nutrient solution with
nitrate as the sole nitrogen source (Hewitt, 1966). Water stress was applied
by addition of polyethylene glycol (PEG) (MW 6000) at 0 (control), 30 and
60 g l-1 for seedling until 14 days and 0, 20 and 40 g l-1
for plants until 28 days after germination. We used PEG 6000 because is
an inert polymer, with a non-ionic and virtually impermeable long chain
(Couper and Eley,1984), which allows to maintain an uniform water potential
during the whole experimental period.
Dry weight
was measured after drying out cotyledons or leaves at 80°C for 72 h.
Chlorophyll
a and b and carotenoids were extracted with acetone (100%)
in a ratio 1:20 (w/v). Pigment concentrations were calculated by Lichtenthaler
method (1987).
Gas exchange
measurements were determined at saturating light intensity, regulated temperature
and relative humidity, using an Infra-red Gas Analyzer (IRGA).
RESULTS

|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
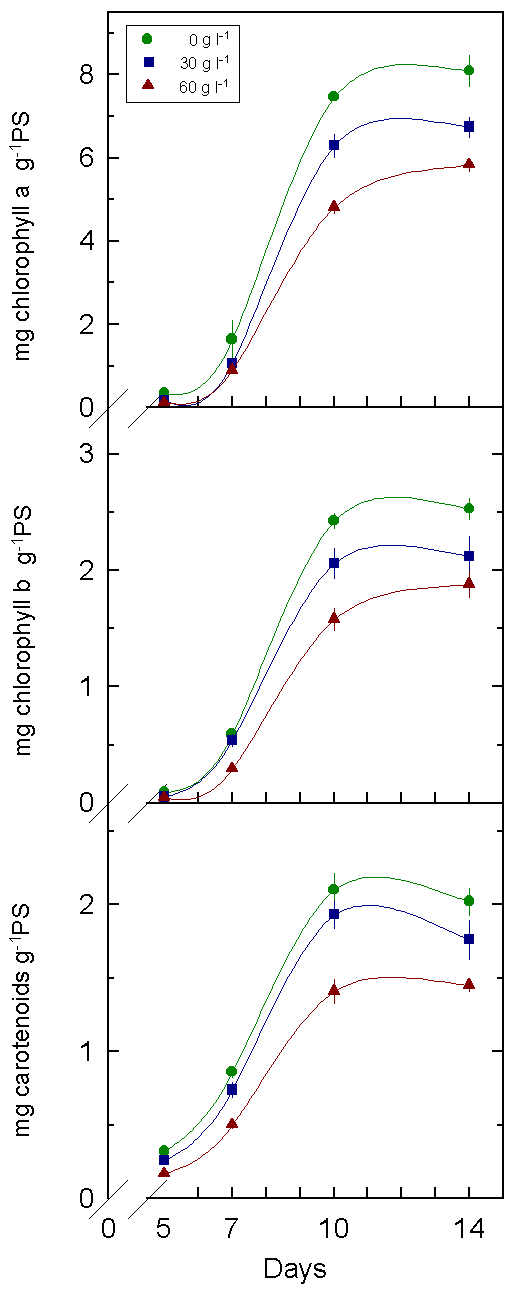
Fig.3: Pigment content
of cotton cotyledons between 5 and 14 days after sowing. Seedlings were
treated with different concentrations of PEG.

|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
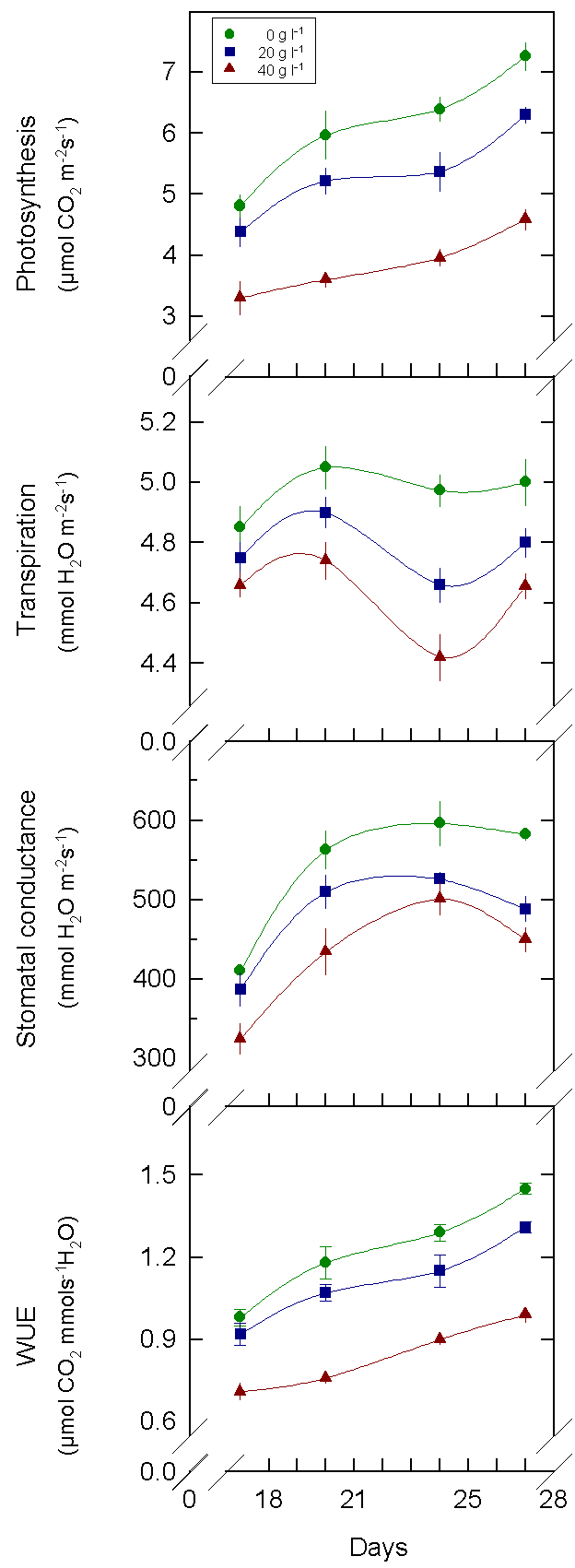
Fig.5. Net photosynthesis,
stomatal conductance, transpiration and water use efficiency of cotton
plants between 18 and 28 days after sowing. Plants were treated with different
concentrations of PEG.
Conclusion
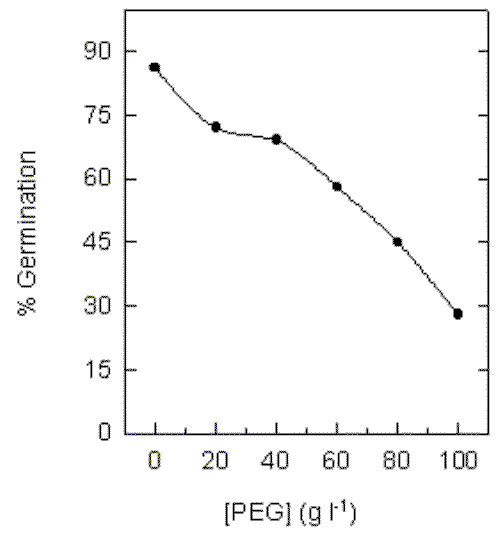
1. Water stress, induced by PEG, causes a decrease in the germination index and in the morphological development of organs from young cotton plants.
2. Dry and fresh weights of cotyledons and fully expanded first leaf were smaller at increasing stress level, which suggests that water absorption and/or retention, as well as biomass gain, are affected by water stress.
3. Water stress diminished the relative growth rate of cotton plants. The net assimilation rate was not significantly decreased, being the major factor responsible for this decrease the leaf area rate.
4. Net photosynthesis, stomatal conductance and transpiration in stressed plants were smaller than in the control, but the photosynthetic rate was more affected than transpiration. Accordingly, water use efficiency (WUE) decreases in PEG-treated plants.
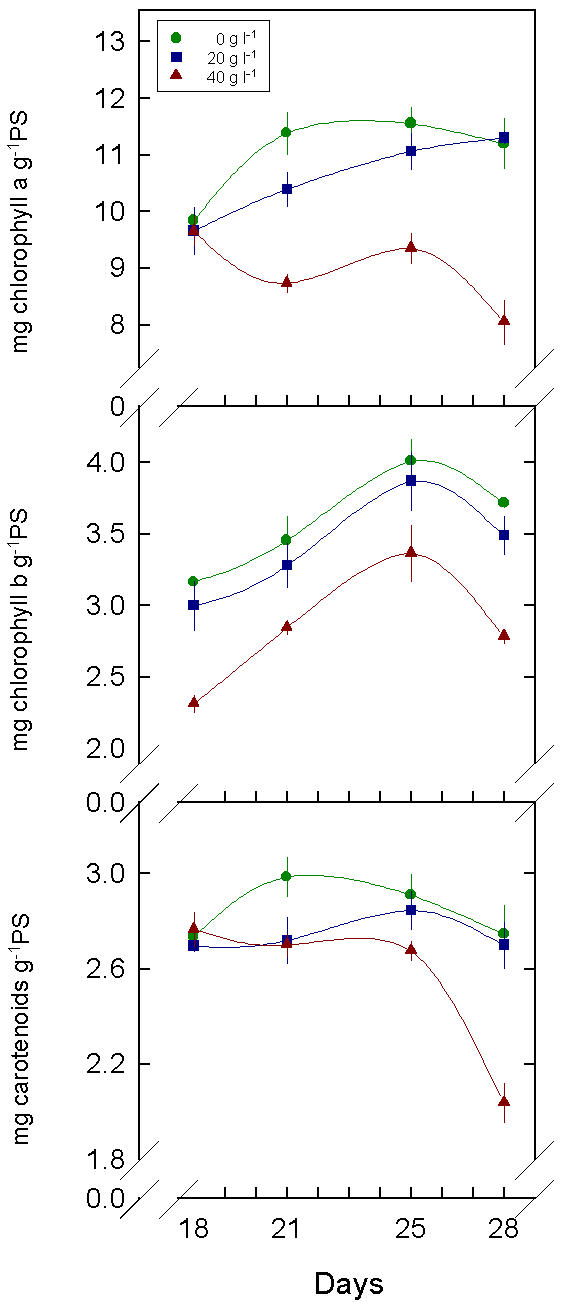
5. The pigment content was smaller in cotyledons and leaves under stress. This fact may explain in part the decrease in the photosynthetic process.
6. The earlier the water stress
is applied, the less successful is the development of cotton plants.
References
1. Couper, A. and Eley,
D. (1948). Surface tension of polyethylene glycol solutions. J. Polymer.
Sci. 3: 345-349.
2. Hewitt, E. J. (1966). Sand
and water culture methods in the study of plant nutrition.
Commonwealth Bureau of Horticultural
and Plantation Crops, East Malling Tech. Commun. 22.
3.Kramer, P.J. and Boyer, J.S.
(1995). Water relations of plants and soils. Academic Press, San Diego.
4.Lichtenthaler, H.K. (1987).
Chlorophylls and carotenoids: Pigments of phothosynthetic biomembranes.
Methods Enzymol. 148: 350-382.
| Discussion Board
| Previous Page
| Your Poster Session
|