Invited Symposium: Iron Transport
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
Millions of people worldwide suffer from nutritional imbalances of essential metals like iron and zinc. These same metals, along with pollutants like cadmium and lead, contaminate soils at many sites around the world. In addition to posing a threat to human health, these metals can poison plants, livestock and wildlife. Deciphering how metals are absorbed, transported, and incorporated as protein cofactors may help solve both of these problems. For example, edible plants could be engineered to serve as better dietary sources of metal nutrients and other plant species could be tailored to remove metal ions from contaminated soils.
We have cloned an iron transporter gene, IRT1, from Arabidopsis thaliana by functional expression in a yeast strain defective for iron uptake. Yeast expressing IRT1 possess a novel Fe(II) uptake activity. IRT1 is predicted to be an integral membrane protein with a potential metal binding domain consisting of the sequence HXHXHXH. IRT1 is expressed only in roots and only in the roots of iron-limited plants. Therefore, we suggest that IRT1 is responsible for iron uptake into the roots of iron-limited plants.
Database analysis and additional genetic and functional cloning analyses has determined that IRT1 is a member of a multigene family of metal ion transporters found in yeast, protozoa, plants, nematodes, and mammals. We refer to this group as the ZIP family of metal ion transporters. The name stands for "ZRT1-, IRT1-like Protein" after the first two members of the family that were characterized in detail. These ZIP proteins define a family of metal ion transporters that are found in a diverse array of organisms making it now possible to address questions of metal ion accumulation and homeostasis in eukaryotes.
Results
SUMMARY OF THE ZIP GENE FAMILY
The known members of the ZIP gene family are described in Table 1. GenBank accession numbers are reported for all submitted sequences. Sequence information for some genes is not complete and their similarity to IRT1 is shown in parentheses. One interesting feature is that Arabidopsis alone contains eight different members of the family. This high number of closely related genes probably reflects genetic redundancy, different substrate specificities, different tissue-specific expression, and perhaps even different subcellular localizations.
Table 1. The members of the ZIP gene family.
----------------------------------------------------------------------
Accession %Similarity Proposed
Gene Organism Number to IRT1 Function
----------------------------------------------------------------------IRT1 A. thaliana U27590 -- Fe uptake
IRT2 " T04324 85 ?
IRT3 " M35868 (68) ?
ZIP1 " AF033535 67 Zn uptake
ZIP2 " AF033536 53 Zn uptake
ZIP3 " AF033537 66 Zn uptake
ZIP4 " U95973 57 Zn uptake
ZIP5 " AC000098 (69) ?
rZIP1 O. sativa D49213 (82) ?
tZIP1 T. brucei AA601838 (51) ?
ZRT1 S. cerevisiae P32804 58 Zn uptake
ZRT2 " X91258 65 Zn uptake
C18A3.2 C. elegans U28944 47 ?
F55F8.9 " U80447 44 ?
C06G8.3 " Z70306 39 ?
F30B5 " U42437 42 ?
mZIP1 mouse AA269515 (48) ?
hZIP1 humans H20615 (43) ?
hZIP2 " Entrez # 998569 53 ?
----------------------------------------------------------------------
Figure 1 describes the amino acid sequence relationships between many of these different transport proteins. It is interesting to note that the plant, fungal, and protosoan genes fall into one subfamily of ZIP transporters whereas the nematode and mammalian genes comprise a second subfamily.
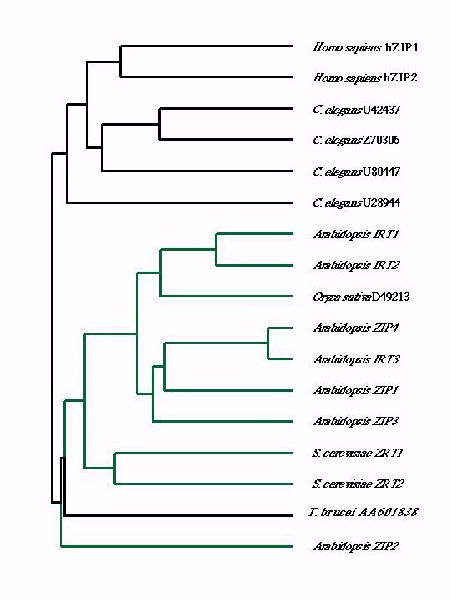 Figure 1. Dendrogram showing the amino acid sequence relationships among ZIP family members.
Figure 1. Dendrogram showing the amino acid sequence relationships among ZIP family members.
Figure 2 describes the predicted membrane topology of most of the members of the family. Most members have eight predicted transmembrane domains and a similar predicted orientation such that the amino and carboxy terminal ends are located on the extracellular surface. These proteins range in size from 309-476 amino acids and this size difference is due to different lengths of the loop region between transmembrane domains III and IV. For this reason, we call this the "variable region". The variable region frequently contains the histidine-rich potential metal binding domain referred to above. Finally, ZIP transporters have no sequence similarity to other families of metal ion transporters including transport ATPases and ABC transporters. Thus, metal ion uptake by these transporters does not appear to be directly linked to ATP hydrolysis.
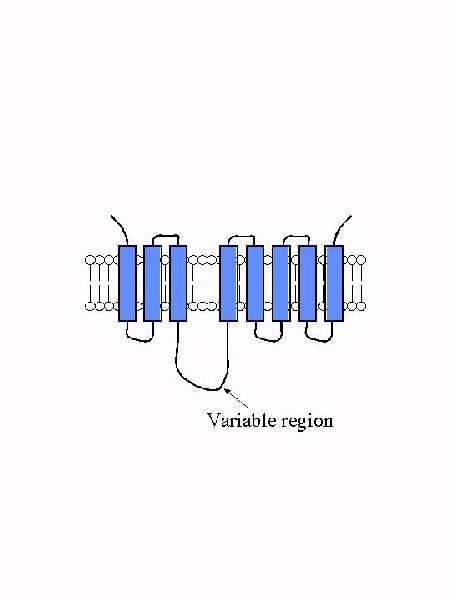 Figure 2. The predicted topology of ZIP transporters.
Figure 2. The predicted topology of ZIP transporters.
CHARACTERIZATION OF THE ZIP ZINC TRANSPORTERS OF ARABIDOPSIS
An underlying hypothesis of our current research is that most, if not all, of the ZIP transporters play roles in the transport of metal ion substrates. Consistent with this hypothesis, we have recently found that four of the ZIP genes from Arabidopsis, ZIP1-4, encode zinc transporters. ZIP1, 2, and 3 were identified by complementation cloning in yeast; expression of these genes suppresses the growth defect of a zinc uptake defective yeast mutant (zrt1 zrt2) cultured on zinc-limiting media (Figure 3). ZIP4 was identified by database analysis through its similarity to other ZIP proteins.
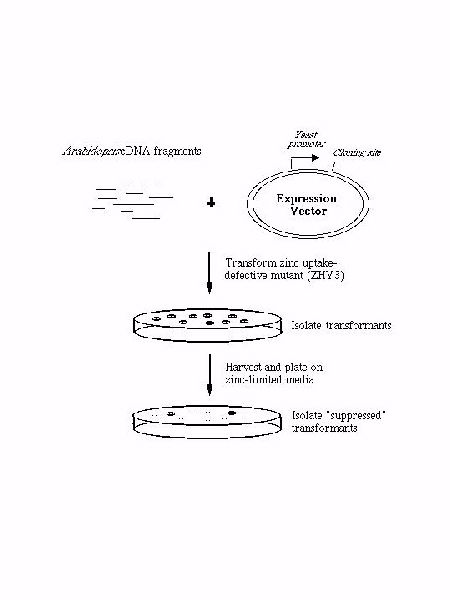 Figure 3. Complementation cloning of Arabidopsis zinc transporters.
Figure 3. Complementation cloning of Arabidopsis zinc transporters.
The ability of ZIP1, ZIP2, and ZIP3 to suppress the growth defect of a zrt1 zrt2 yeast mutant suggested that these genes may encode zinc transporters. To test this hypothesis, we examined 65Zn accumulation in a zrt1 zrt2 strain expressing each of these genes. At 0o C, no zinc accumulation was detected in any of the strains tested or in an untransformed control strain (Fig. 4A and B).
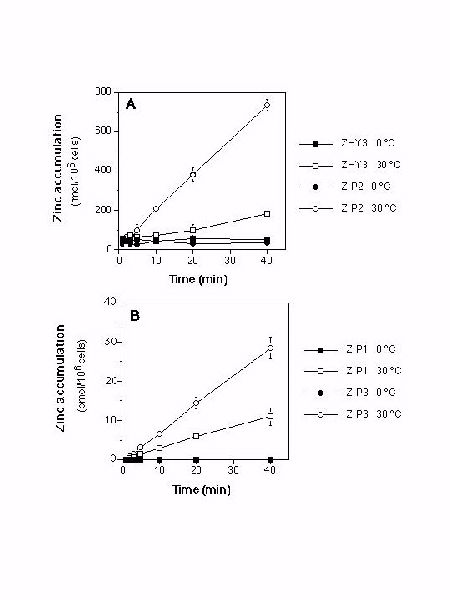 Figure 4. Zinc accumulation over time was assayed in exponentially growing untransformed zrt1 zrt2 yeast cells (ZHY3) or ZHY3 cells expressing ZIP1, ZIP2, ZIP3. Accumulation was assayed at 0o C and 30o C and at 1 (ZIP2) or 10 uM (ZHY3, ZIP1, ZIP3) 65Zn.
Figure 4. Zinc accumulation over time was assayed in exponentially growing untransformed zrt1 zrt2 yeast cells (ZHY3) or ZHY3 cells expressing ZIP1, ZIP2, ZIP3. Accumulation was assayed at 0o C and 30o C and at 1 (ZIP2) or 10 uM (ZHY3, ZIP1, ZIP3) 65Zn.
At 30o C, no zinc accumulation was detected in the untransformed control strain whereas a high rate of accumulation was observed in the ZIP1- and ZIP3-expressing strains (approximately 270 and 710 fmol/min/106 cells, respectively) (Figure 4B). The rate of ZIP1- or ZIP3-dependent zinc accumulation was not greatly altered by variations in pH of the assay solution between 4.0 and 6.0 (data not shown). In contrast, ZIP2-dependent zinc accumulation was highly pH-dependent; no uptake activity was detectable in the ZIP2-expressing strain when assayed at pH values below 5.0 (data not shown) yet at pH 6.0, temperature-dependent zinc accumulation was observed (approximately 19 fmol/min/106 cells) (Figure 4A). A cDNA version of the ZIP4 coding sequence, constructed using PCR, was cloned into pFL61 and used in yeast complementation and uptake assays. No uptake could be detected in yeast cells expressing the ZIP4 gene nor could ZIP4 complement the growth defect of the zrt1 zrt2 mutant on zinc-limited medium. This may be due to subcellular rather than plasma membrane localization of ZIP4 in yeast.
When assayed over a range of zinc concentrations, uptake activity dependent on expression of the ZIP1, ZIP2, and ZIP3 genes was concentration-dependent and saturable (Fig. 5).
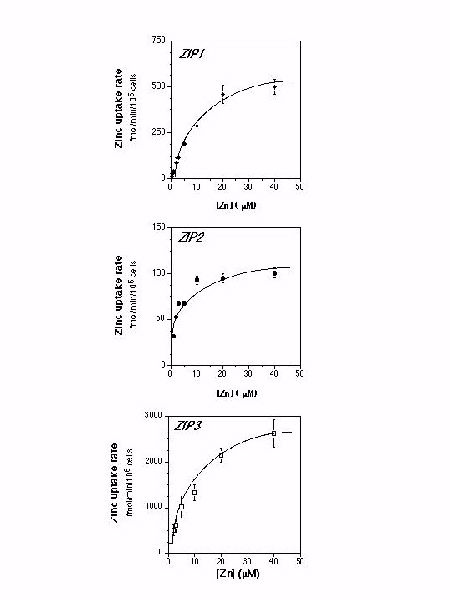 Figure 5. Concentration dependence of zinc uptake by ZIP1, 2, and 3. The initial rate of zinc uptake was assayed in exponentially growing yeast cells (ZHY3) expressing ZIP1, ZIP2, or ZIP3 over a range of 65Zn concentrations. The Michaelis-Menten kinetic values derived from these data are described in Table 2. Interestingly, the apparent Kmvalues of these transporters are similar to the levels of zinc commonly found in plant rhizospheres.
Figure 5. Concentration dependence of zinc uptake by ZIP1, 2, and 3. The initial rate of zinc uptake was assayed in exponentially growing yeast cells (ZHY3) expressing ZIP1, ZIP2, or ZIP3 over a range of 65Zn concentrations. The Michaelis-Menten kinetic values derived from these data are described in Table 2. Interestingly, the apparent Kmvalues of these transporters are similar to the levels of zinc commonly found in plant rhizospheres.
Table 2. Michaelis-Menten kinetic constants of zinc uptake in yeast. These values are estimated from the data in Figure 5.
----------------------------------------------------------- Gene Apparent Km (uM) Vmax (fmol/min/106 cells) ----------------------------------------------------------- ZIP1 13 � 2 693 � 43 ZIP2 2 � 0.3 107 � 4 ZIP3 14 � 2 3528 � 207 -----------------------------------------------------------
To assess if metals in addition to zinc are substrates for ZIP1, ZIP2, or ZIP3, we tested other metal ions for their ability to inhibit zinc uptake mediated by these proteins.
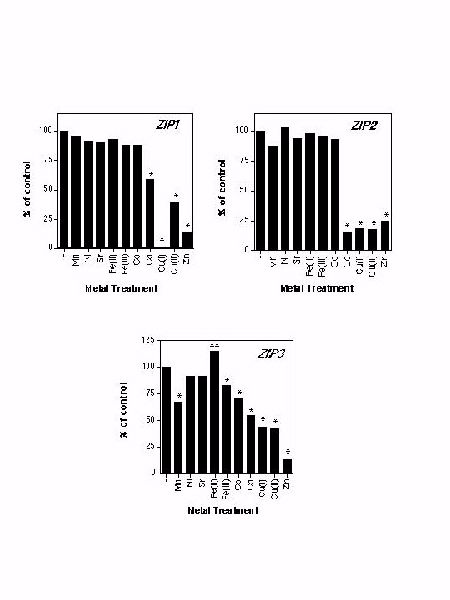 Figure 6. Assessing the substrate specificity of ZIP1, ZIP2, and ZIP3 for zinc. ZHY3 cells expressing the indicated gene were assayed for their zinc uptake rate with 10 (ZIP1, ZIP3) or 1 uM (ZIP2) 65Zn in the absence (-) or presence of a 10-fold excess of the indicated metal ions. The asterisks represent significant differences from control values (P<0.05) as determined by one-way ANOVA.
Figure 6. Assessing the substrate specificity of ZIP1, ZIP2, and ZIP3 for zinc. ZHY3 cells expressing the indicated gene were assayed for their zinc uptake rate with 10 (ZIP1, ZIP3) or 1 uM (ZIP2) 65Zn in the absence (-) or presence of a 10-fold excess of the indicated metal ions. The asterisks represent significant differences from control values (P<0.05) as determined by one-way ANOVA.
Zinc uptake by ZIP1 was not inhibited by a 10-fold excess of Mn, Ni, Fe, or Co (Figure 6). Zinc was the most potent competitor demonstrating that ZIP1 prefers zinc as its substrate over these other metal ions. Cd and Cu also inhibited zinc uptake but to a lesser extent. Although the mechanism of these inhibitory effects is not yet known, these results suggest that Cd and Cu may also be substrates for ZIP1. Zinc was the most potent competitor demonstrating that ZIP1 prefers zinc as its substrate over these other metal ions. Zinc uptake mediated by ZIP3 was inhibited by Mn, Fe, Co, Cd, and Cu (Fig. 6). Again, zinc had the greatest effect indicating that ZIP3 also prefers zinc. Cd and Cu inhibited zinc uptake mediated by ZIP2 to as great a degree as zinc (Fig. 6) suggesting that ZIP2 may also have a high affinity for Cd and/or Cu. In summary, these three transporters show unique sensitivities to other metal ions which may reflect differences in their substrate specificities. We have found that expression of ZIP1, ZIP2, ZIP3 and ZIP4 in a yeast strain defective for iron uptake (a fet3 fet4 mutant, DEY1453) failed to alter this strain�s iron-limited growth defect or its iron uptake activity. Thus, the ZIP proteins cannot transport iron.
Why does Arabidopsis have multiple zinc transporters? Zinc, like other metal ions, is transported from the soil into the root and then must cross both cellular and organellar membranes as it is distributed throughout the plant. Specific zinc transporters may play different roles in this process. Northern blot analysis shown in Figure 7 demonstrated that ZIP1 and ZIP3 are zinc responsive.
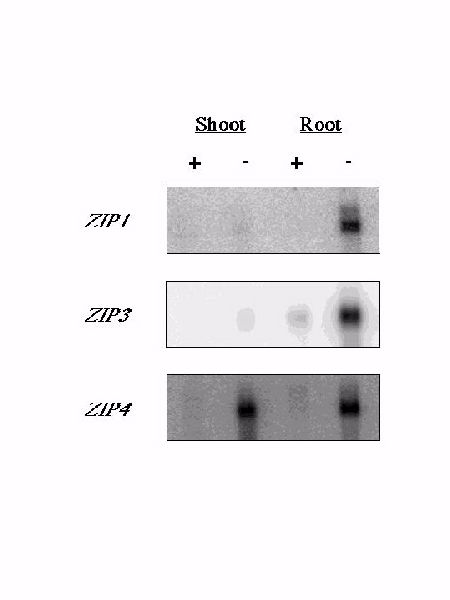 Figure 7. Gene-specific probes were hybridized to a Northern blot containing 1 ug of polyA+ mRNA prepared from the roots or shoots of hydroponically grown zinc-sufficient (+) or zinc-deficient (-) plants. Expression of ZIP2 has not been detected despite repeated attempts.
Figure 7. Gene-specific probes were hybridized to a Northern blot containing 1 ug of polyA+ mRNA prepared from the roots or shoots of hydroponically grown zinc-sufficient (+) or zinc-deficient (-) plants. Expression of ZIP2 has not been detected despite repeated attempts.
ZIP1 and ZIP3 are expressed predominantly in the roots of zinc-deficient plants; little or no mRNA was detected in the roots of zinc-sufficient plants or in the shoots of zinc-sufficient or zinc-deficient plants (Figure 7). Despite repeated attempts, ZIP2 mRNA could not be detected in plants grown under either condition. ZIP4 also responds to zinc deficiency but unlike ZIP1 and ZIP3, this gene is induced in both the shoots and roots of zinc-deficient plants. This result is especially interesting given that sequence analysis algorithms predict that ZIP4 is localized to chloroplasts (see below). The results of our Northern blot analyses are consistent with a role for ZIP1 and ZIP3 in the uptake of zinc from the rhizosphere and a role for ZIP4 in the transport of zinc in plastids. It is not surprising that ZIP gene expression would be regulated; zinc is toxic when present in excess. We also examined ZIP gene expression in iron-sufficient and iron-deficient plants and saw no response of any of the ZIP genes to the same conditions of iron-deficiency that induce IRT1 expression (data not shown). Thus, ZIP1, ZIP3, and ZIP4 respond specifically to a lack of zinc.
Discussion and Conclusion
The predicted products of the ZIP1, ZIP2, ZIP3 and ZIP4 genes are 355, 353, 339 and 374 amino acids in length, respectively. As with many other ZIP family members, these proteins contain eight potential transmembrane domains. ZIP1, ZIP2, and ZIP3 each have a potential signal sequence and are predicted to be plasma membrane proteins; ZIP4 contains a potential chloroplast targeting sequence [PSORT]. The potential metal binding motif, containing multiple histidine residues, is found in the variable regions of almost all of the fully-sequenced members of this family including IRT1, IRT2, ZIP1, ZIP4, ZRT1 and ZRT2. In contrast, ZIP2 and ZIP3 contain only a single histidine in this region. Studies of ZRT1 suggest that the histidine-rich motif is located on the intracellular face of the plasma membrane and also indicate that it is essential for transporter function (data not shown). Three of the transmembrane domains (II, IV and V) contain a histidine residue that is fully conserved among all family members. These histidines are predicted to lie on the polar face of amphipathic helices, suggesting a possible role for these residues in substrate transport through the membrane. Furthermore, conserved acidic amino acids present in the transmembrane domains of several of the ZIP proteins may also be important for substrate movement.
The ZIP genes of Arabidopsis represent four new members of a rapidly growing family of eukaryotic proteins. To date, seven of the family members have been implicated in metal ion transport. These results demonstrate that the ZIP transporter family plays roles in metal ion metabolism in a diverse array of eukaryotic organisms. The ZIP family is structurally distinct from other metal ion transporters such as the CDF family that includes the recently identified mammalian zinc effluxers, P-type ATPases that are involved in the transport of a variety of cations, and the Nramp proteins recently implicated in divalent cation transport. The ZIP genes then offer a good starting point for understanding how metals cross membranes.
Our main conclusions are as follows.
1. The ZIP genes encode a family of metal ion transport proteins found in a diverse array of eukaryotic organisms. No members have yet been identified in bacteria. This may mean that ZIP transporters arose in the evolution of eukaryotes or, more likely, prokaryotic relatives have diverged sufficiently from the eukaryotic proteins to be unrecognizable.
2. Expression of ZIP1, ZIP2, and ZIP3 in yeast suppresses the zinc-limited growth defect of a mutant defective for zinc uptake.
3. ZIP1, ZIP2, and ZIP3 are functional zinc transporters when expressed in yeast. ZIP4-dependent zinc uptake in yeast was not detected.
4. The apparent Km values for ZIP1, ZIP2, and ZIP3 (2-14 uM) are similar to those measured in whole plant roots. These results suggest that these transporters are of sufficiently high affinity to be physiologically relevant.
5. Zinc uptake by the ZIP transporters in yeast is sensitive to inhibition by some other metal ions suggesting differences in their substrate specificities. Further studies will be required to fully address this question.
6. ZIP1, ZIP3, and ZIP4 expression is induced under zinc-limiting conditions. These results support the hypothesis that these genes play a role in zinc uptake in the plant. ZIP1 and ZIP3 are expressed primarily in the root and may be responsible for acquisition of zinc from the soil. ZIP4 is expressed in both roots and shoots suggesting that this gene may play roles in zinc transport elsewhere in the plant.
ACKNOWLEDGMENTS
This work was supported by grants from the NIH, NSF, and DOE to D.E. and M.L.G.
References
- Zhao, H. and Eide, D. The yeast ZRT1 gene encodes the zinc transporter protein of a high affinity uptake system induced by zinc limitation. Proc. Natl. Acad. Sci. USA 93:2454-2458 (1996).
- Eide, D., Broderius, M., Fett, J., and Guerinot, M.L. A novel, iron-regulated metal transporter from plants identified by functional expression in yeast. Proc. Natl. Acad. Sci. USA 93:5624-5628 (1996).
- Zhao, H. and Eide, D. The ZRT2 gene encodes the low affinity zinc transporter in Saccharomyces cerevisiae. J. Biol. Chem. 271:23203-23210 (1996).
- Grotz, N., Fox, T., Connolly, E., Park, W., Guerinot, M.L., and Eide, D. Identification of a family of zinc transporter genes from Arabidopsis thaliana that respond to zinc deficiency. Proc. Natl. Acad. Sci. USA 95:7220-7225 (1998).
| Discussion Board | Previous Page | Your Symposium |