Invited Symposium
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Results
Model Chamber for Perturbed Flow (PF): To study the effects PF on VCAM-1 expression, HAEC-lined circular cover slips were "glued" into the asymetrically placed grooves in the chamber, thus exposing the cells at distinct local flow patterns of pulsatile/perturbed flow: Cover slips 1, 3, and 5 are in the path of the pulsatile jet-stream, while cover slips 2 and 4 are located in the areas of low-shear, recirculating eddies which closely reflect the PF patterns found at sites of atherosclerotic predilection in vivo. Shown in Figure 1 is a flow-visualization analysis of the various flow patterns in the culture chamber with the inserted coverslips.
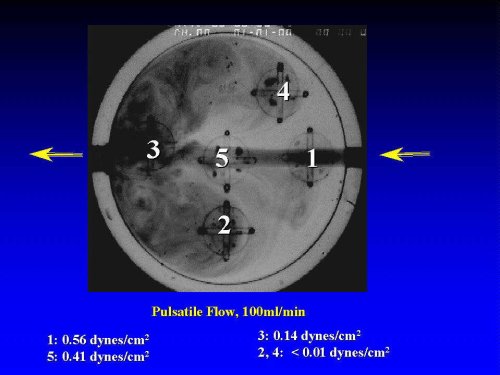 Figure 1: Macroscopic View of the Circular Flow Chamber. Five microscope cover slips (covered with a confluent monolayer of HAEC) are placed asymmetrically into the chamber, Flow visualization analysis, based on dye -injection, demonstrates the distinct local flow patterns, including jet stream (positions 1,3,5) and circulating eddies (perturbed flow, positions 2,4). The shear stress is varied by adjusting the flow rate, and/or the height of the flow chamber. .
Figure 1: Macroscopic View of the Circular Flow Chamber. Five microscope cover slips (covered with a confluent monolayer of HAEC) are placed asymmetrically into the chamber, Flow visualization analysis, based on dye -injection, demonstrates the distinct local flow patterns, including jet stream (positions 1,3,5) and circulating eddies (perturbed flow, positions 2,4). The shear stress is varied by adjusting the flow rate, and/or the height of the flow chamber. .
Quantitation of Perturbed Flow-induced VCAM-Protein Expression by ELISA: We recently reported that under static conditions, the base-line levels of VCAM-1 in HAEC were very low and upregulated up to 40 fold by TNF (Kanda et al.,1998). Upon exposure to pulsatile flow, VCAM-1 protein levels were upregulated by the various types of low-shear (0.1 - 0.56 dyne/cm2), pulsatile flow in a time- and flow-pattern dependent fashion, reaching a peak after ca. 5 h which was maintained for at least 24 h (not show n). For example, VCAM-1 levels in HAEC placed in position 2 and 4, representing the areas of PF, were elevated within 5 h by approximately 7-8 fold over base-line and persisted over 24 hours of continuous flow (Figure 2). By comparison, exposure to the pulsatile jet stream (in positions 1,3,5) yielded qualitatively similar, yet smaller increases in VCAM-1 expression.
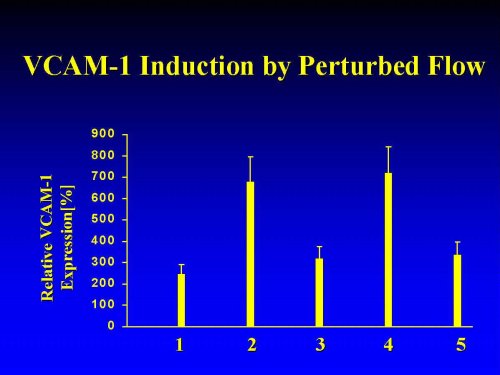 Figure 2: Positional Dependence of Low Shear-Induced VCAM-1 Protein Expression. VCAM-1 expression in HAEC exposed for 8 h to pulsatile flow was measured by fluorescent ELISA, as previously described (Kanda et al., 1998). All data are normalized to basal VCAM-1 expression in the untreated static controls and expressed as means � SEM (n=8).
Figure 2: Positional Dependence of Low Shear-Induced VCAM-1 Protein Expression. VCAM-1 expression in HAEC exposed for 8 h to pulsatile flow was measured by fluorescent ELISA, as previously described (Kanda et al., 1998). All data are normalized to basal VCAM-1 expression in the untreated static controls and expressed as means � SEM (n=8).
The positional differences in pulsatile flow-induced VCAM-1 upregulation between cells located, respectively, on cover slips 2,4 and cover slips 1,3,5 might reflect differential EC activation by distinct local flows-patterns, viz., recirculating eddies/PF vs. jet stream in positions 2,4 and 1,3,5 , respectively. On the other hand, the shear stress values in these areas differ: For a flow rate of 100 ml/min, local shear stress values were calculated to amount to 0.1 dynes/cm2 in positions 2,4 and to 0.56 dynes/cm2 in positions 1,3,5 (Christensen et al., 1992; Samet and Lelkes, 1993, 1999). Thus, the differences might reflect a sensitivity of VCAM-1 to low shear stress of diverse magnitudes. Past studies suggest that threshold values of 2-5 dynes/cm2 are required for the induction shear stress-sensitive phenomena by steady, laminar flow. By contrast, we previously reported physiological consequences of pulsatile flow-induced shear stress at these low magnitudes , such as morphological changes in endothelial cells (Lelkes and Samet, 1991; Samet et al., 1993).
In studies, to be reported elsewhere, we established that PF-induced VCAM-1 protein expression is a transcriptional/translational event, which is inhibited by preincubating the cells with either actinomycin D or cycloheximide. Moreover, exposure to PF leads to an upregulation of VCAM-1 steady-state mRNA levels (Lelkes et al., manuscript in preparation)
In order to test the dose/shear-stress responsiveness of VCAM-1 protein induction by PF, we increased the fluid shear stress approximately 2 fold without altering the flow characteristics, by reducing the thickness of the flow chamber to �.
Table I
Effect of Non-Laminar Shear Stress on VCAM-1 Expression in HAEC
----------------------------------------------------------------------- Recirculating Eddies(positions 2/4) Pulsatile Jet(positions 1/3/5) ----------------------------------- ------------------------------ 0.1 dynes/cm2 0.2 dynes/cm2 0.56 dynes/cm2 1.1 dynes/cm2 ----------------------------------------------------------------------- 766 � 123 953�207 347�152* 308 � 85* ------------------------------------------------------------------------ All values (means � SEM) are normalized to VCAM-1 expression under static conditions. * p < 0.05 by comparison to 0.1 dynes/cm2.
As seen in Table I , exposure for18 h to PF with a shear stress of approximately 0.2 dynes/cm2, resulted in a trend (statistically not significant, p > 0.05) to increase VCAM-1 expression in cells positioned in the recirculating eddies. By contrast, we failed to obtain statistically significant alterations in the PF-induced VCAM-1 expression in cells in positions 1,3,5, in which the shear stress in the jet was elevated up to approx. 1.1 dynes/cm2 .
Inhibitory Effect of PDTC: Cytokine-induced VCAM-1 expression is mediated via activation of the reactive oxygen species(ROS) sensitive nuclear transcription Factor NF-kappaB (Collins et al., 1995). In preliminary studies we established that exposure of confluent endothelial cell monolayers for up to 8 hours to the ROS scavenger and putative NF-kappaB inhibitor pyrollidine dithiocarbamate (PDTC, 10 �M), reduced basal VCAM-1 expression. Moreover, when the cells were first preincubated for 30 min with PDTC and then subsequently stimulated for 8 h with 1ng/ml TNF, the cytokine-induced upregulation of VCAM-1 was completely abrogated (not shown).
In preliminary studies we established the dose response of PDTC-inhibition of PF-induced VCAM-1 upregulation. For the sake of simplicity, we initially focused on the cells located in the recirculating eddies (positions 2 and 4). To test the dose response of PDTC, the cells were preincubated for 30 min with 0.001 to 100 �M of PDTC and then exposed for 5 hr s to PF. The ID50 for PDTC inhibition was approx. 0.0.1 �M. Flow-induced VCAM-upregulation was essentially abrogated at PDTC doses exceeding 3 �M (not shown). Therefore all subsequent studies were carried out with 10 �M PDTC. When the cells were preincuabted for 30 min with 10 �M PDTC and then, in the continuous presence of 10 �M PDTC, exposed for 8 h to pulsatile flow, upregulation of VCAM-1 expression was largely abrogated. The extent of attenuation/ abrogation of the flow-effects of VCAM-1 expression was independent of the local flow pattern and/or of the level of fluid shear stress (Figure 3). In preliminary studies, using other antioxidants, such as N-acetyl cysteine, we obtained comparable results (not shown).
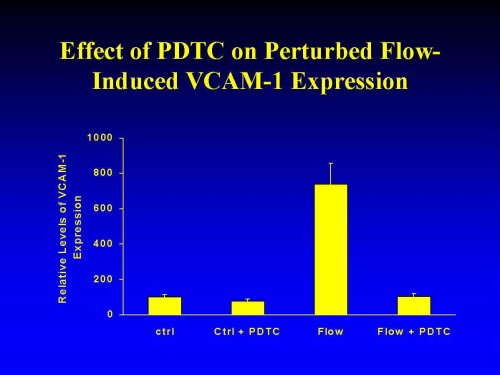 Figure 3: Inhibition of Flow-Induced VCAM-1 Expression by pyrollidine dithiocarbamate (PDTC). Circular cover slips lined with confluent monolayers of HAEC were mounted into the flow chambers, incubated for 30 min with 10 �M pyrollidine dithiocarbamate, and then exposed for 8 h to pulsatile flow. VCAM-1 expression was assessed by ELISA. For the sake of clarity only pooled values of VCAM-1 expression in cells from cover slips 2/4 are shown. All data are normalized to basal VCAM-1 expression in the untreated static controls and expressed as means � SEM (n=3).
Figure 3: Inhibition of Flow-Induced VCAM-1 Expression by pyrollidine dithiocarbamate (PDTC). Circular cover slips lined with confluent monolayers of HAEC were mounted into the flow chambers, incubated for 30 min with 10 �M pyrollidine dithiocarbamate, and then exposed for 8 h to pulsatile flow. VCAM-1 expression was assessed by ELISA. For the sake of clarity only pooled values of VCAM-1 expression in cells from cover slips 2/4 are shown. All data are normalized to basal VCAM-1 expression in the untreated static controls and expressed as means � SEM (n=3).
To determine the temporal window, in which PDTC can abrogate pulsatile flow-induced VCAM-1 upregulation, we either pre-incubated HAEC with PDTC (10 mM), or added the drug at various time points during the flow studies. VCAM-1 expression was measured after 4.5 h exposure to flow. Inhibition of VCAM-1 induction by PDTC was fully effective if the drug was added to the experiment either prior to, concomitantly with, or up to 90 minutes after switching on the flow. However, when added at 180 minutes after onset of the flow or later, PDTC was unable to inhibit the induction of VCAM-1 (not shown). These results suggest that the PDTC-sensitive cellular/ molecular events, which are activated upon exposure of the cells to PF, might represent some of the immediate/early signaling steps Potential candidates might include flow-activated second messenger/signaling pathways, such as ROS and ROS-sensitive transcription factors.
Signaling Mechanisms: Taken together, the above results suggest that PF activation of HA EC initiates a cascade of cellular events which results in the enhanced expression of VCAM-1 protein levels. We hypothesized that upregulation of VCAM-1 by PF might involve specific tyrosine phosphorylation- dependent intracellular signaling cascades. To test our hypothesis we tested the effects of PF on the activation of specific signaling proteins, such as MAPK (erk-1) and FAK, as well as the nuclear translocation of several, putative, shear-stress responsive transcription factors NF-kappaB, AP-1 SP1 and egr-1.
Preliminary analysis of whole cell lysates of PF-activated HAEC by Western Blotting using a monoclonal anti phosphotyrosine antibody 4G10 revealed numerous phosphorylated bands, including bands at approx. 40 and 125 kD characteristic of activation of erk (extracellular receptor activated kinase) and FAK (focal adhesion kinase). Tyrosine phosphorylation of many of these bands, including pp40 and pp125 was inhibited by pretreating the cells with 10 �M PDTC (not shown).
Previous studies suggested that laminar flow of high shear activated mitogen activated protein kinase (MAPK)- dependent cellular signaling pathways (Ishida et al., 1996). To assess the effects of low shear PF on MAPK signaling, we evaluated the activation (protein tyrosine) phosphorylation by immunoprecipitation from HAEC exposed for 15 min to pulsatile flow. As seen in Figure 4, erk was activated by PF in a manner which appeared to be independent of the particular local flow patterns. Similarly, no positional variation was detected in PF-activated FAK . Therefore, we conclude, that neither the integrin/FAK nor the MAPK kinase pathways are suitable candidates for a unique mechanisms by which EC might sense perturbed flow patterns, leading to VCAM-1 upregulation.
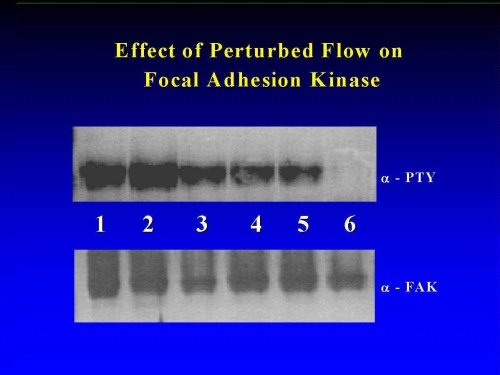 Figure 4: Tyrosine Phosphorylation of erk and of FAK in PF-Activated HAEC. Confluent monolayers of HAEC (n =4) were exposed for 15 min to pulsatile flow. Erk and FAK were immunoprecipitated with monospecific polyclonal antibodies (from Santa Cruz) and analyzed by Western blotting for activation (tyrosine phosphorylation, -PTY) and for quantification (-erk, -FAK). Lanes 1-5: corresponding positions 1-5 in the flow chamber, lane 6: static controls. For details, see Materials and Methods.
Figure 4: Tyrosine Phosphorylation of erk and of FAK in PF-Activated HAEC. Confluent monolayers of HAEC (n =4) were exposed for 15 min to pulsatile flow. Erk and FAK were immunoprecipitated with monospecific polyclonal antibodies (from Santa Cruz) and analyzed by Western blotting for activation (tyrosine phosphorylation, -PTY) and for quantification (-erk, -FAK). Lanes 1-5: corresponding positions 1-5 in the flow chamber, lane 6: static controls. For details, see Materials and Methods.
Gene-activation by laminar shear stress is preceded by the nuclear translocation of specific transcription factors. In recent years several such shear-responsive transcription factors have been identified, such as AP-1, NF-kappaB and SP1 (Lan et al., 1994; Lin et al., 1997). Kachigian and coworkers recently demonstrated that NF-kappaB bind sites correspond to nucleotide sequences which are also contained in the "classical" shear stress response element (Khachigian et al., 1995). Using the electrophoretic mobility shift assay, exemplified for potion 2/4 we found that PF failed to activate the ROS- inducible nuclear transcription factor NF-kappaB (Figure 5). A similar negative result was also obtained in positions 1/3/5 (not shown). This finding is surprising, since, as shown above, VCAM-1 induction by PF is abrogated by the ROS antagonists PDTC. Indeed, additional activation of the cells during the flow experiments with TNF (1 ng/ml) resulted in a significant upregulation of NF-kappaB. Thus, this transcription factor, which previously were identified as being centrasl for the "activation" of certain flow-responsive genes by laminar shear stress, exhibited a rather "unorthodox" responses upon exposure of the cells to PF.
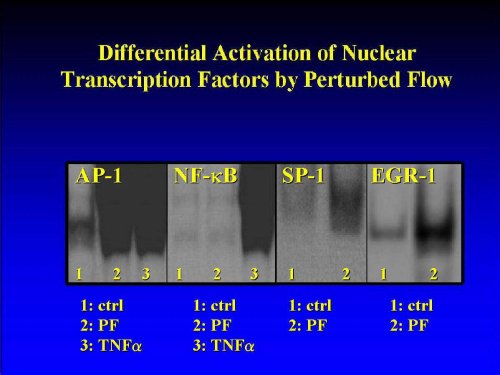 Figure 5: Differential Induction of Nuclear Transcription Factors by Effect of Pulsatile Flow: The nuclear translocation (activation) of transcription factors NF-kappaB, AP-1, egr-1 and Sp1 from HAEC exposed to 30 min of pulsatile flow was assessed using the electrophoretic mobility shifty assay (EMSA), as detailed in Materials and Methods.
Figure 5: Differential Induction of Nuclear Transcription Factors by Effect of Pulsatile Flow: The nuclear translocation (activation) of transcription factors NF-kappaB, AP-1, egr-1 and Sp1 from HAEC exposed to 30 min of pulsatile flow was assessed using the electrophoretic mobility shifty assay (EMSA), as detailed in Materials and Methods.
By contrast, other, more recently discovered shear-responsive transcription factors, such as activator protein 1 (AP-1) (Lan et al., 1994), early growth response gene 1 (egr-1 (Khachigian et al., 1994, 1995, 1996) and SP-1 were induced by PF (Figure 5), making these molecules candidate transcription factors involved in the pulsatile-flow induced signalization process, which leads to the upregulation of VCAM-1.Preliminary results suggest a distinct pattern of regulation of transcription factors by the local flow patterns (Lelkes et al., manuscript in preparation).
| <= Materials & Methods | RESULTS | Discussion & Conclussions => |
| Discussion Board | Next Page | Your Symposium |