Neuroscience Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
PLG is an endogenously derived hypothalamic factor originally identified as melanocyte stimulating hormone release inhibiting factor (MIF-1)9,11 and was soon discovered to have an effect on central dopaminergic neurotransmission4,11. Of particular interest to us is the manner in which PLG exerts an effect on the function of dopamine D2-receptors in the basal ganglia. The modulatory effects of PLG on D2-dopaminergic neurotransmission has been demonstrated in a number of in vivo and in vitro studies. In vivo studies have shown that PLG potentiates the behavioral effects of dopamine agonists (enhanced rotation in the hemi-Parkinson's rat and enhanced arousal in Parkinsonian patients)4,12-19, attenuates neuroleptic and morphine induced catalepsy and dopamine receptor supersensitivity4,5, antagonizes oxotremorine induced tremor20-23, potentiates CPP (NMDA receptor antagonist) induced locomotion and darting and protects against MPTP induced degeneration of nigrostriatal dopaminergic neurons24. In vitro radioligand binding studies have shown that PLG increases the binding of D2 agonists in striatal membranes4,11,25-31, modulates the high affinity state of D2 receptors7,11,32 and attenuates neuroleptic-induced D2 receptor supersensitivity1-3.
Dopaminergic neurotransmission in the basal ganglia plays an important role in psychomotor processes. Disruptions or alterations in the function of central dopaminergic pathways contributes to diseases such as Parkinson's and schizophrenia. The ability of PLG to modulate the function of central dopaminergic pathways indicates a potential therapeutic benefit in the treatment of Parkinson's and schizophrenia12-16,33.
RATIONALE.
PLG is an endogenous neuropeptide with known effects on numerous neurological processes. Prolonged PLG treatment has known effects on central dopaminergic neurotransmission. It is reasonable to expect that a long-term elevation of PLG levels will alter the expression of proteins involved in mediating its effects (particularly in the striatum) as happens in other neurotransmitter systems34. Furthermore, observations in our lab indicate that PLG has the ability to modulate the expression of immediate early genes35. ddPCR is a technique that enables the detection of numerous genes that are differentially expressed in two or more tissues. Therefore, ddPCR can be used to identify genes that are differentially regulated by prolonged PLG treatment. Genes found to be regulated by PLG will likely be important modulators of its effects and may provide insight into the mechanism of action of PLG.
HYPOTHESIS.
The hypothesis I tested is that a protracted treatment with PLG alters the expression of genes directly involved in PLG signal transduction. Through the use of ddPCR, I have identified a number of genes whose expression is altered following PLG treatment. It is hoped that this strategy will allow for the identification of genes directly involved in mediating the effects of PLG.
Materials and Methods
Animal Treatment.
Two groups of animals (10 animals per group) were treated daily for 21 days with PLG (20 mg/kg, i.p.) or saline. Following a 2 day washout period, the animals were sacrificed, their brains removed and placed on ice for dissection. Various regions of the brain (striatum, pre-frontal cortex, medulla, cerebellum, hypothalamus, olfactory tubercle and hippocampus) are dissected out and immediately frozen on dry ice. The samples are stored at -80°C until needed.
RNA Preparation.
Total RNA from the striatum is isolated using the Tripure reagent method (Gibco Life Technologies, Gaithersburg, MD). The amount of mRNA obtained was determined by measuring the OD at 260 nm and the purity by the ratio of 260/280 nm. The integrity of the RNA (18S and 28S bands) was assessed by running 5 µg on a denaturing 1% agarose gel. Prior to use in RT-PCR, mRNA is purified using a DNase I (Gibco BRL Life Technologies) treatment to remove any contaminating chromosomal DNA. DNase treatment is done immediately prior to RT-PCR to ensure that the diluted mRNA is not degraded during storage.
Differential Display PCR.
Differential display RT-PCR (ddPCR) was performed using the GeneHunter (Nashville, TN) ddPCR RNAimage kit #1 (figure 1). Briefly, the purified mRNA undergoes three reverse transcription reactions with three different oligo-dT primers (H-T11G, H-T11C, H-T11A). The products of each RT reaction are used in 8 different PCR reactions (MJ Research MiniCycler). Each PCR reaction uses the same oligo-dT primer used in RT and one of eight arbitrary primers. Hence, each mRNA sample undergoes a total of 24 PCR reactions.
Acrylamide Sequencing Gel.
For comparisons between control and PLG treated samples, the ddPCR process is conducted side by side on equal amounts of mRNA from both samples. Following the completion of the ddPCR reactions, the PCR products were separated by running the samples through a 6% denaturing acrylamide gel using a BioRad Sequi-Gen GT nucleic acid electrophoresis cell. Following electrophoresis, the gel is removed from the gel apparatus and dried using a BioRad gel slab drier. The dried gel is exposed to Kodak BioMax MR film for 3 days at room temperature. PCR products from similar reactions for the control and PLG samples are run in adjacent lanes for easy comparison of amplified bands. Those bands identified as being regulated by PLG were excised from the gel and solubilized in 100 µl dH2O, precipitated and finally resuspended in 10 µl dH2O.
cDNA Cloning and Sequencing.
The recovered cDNA (PCR fragments) is reamplified using the same primers by which they were generated. The resultant PCR products are run on a 1.7% agarose gel and their size is checked with what is expected from the acrylamide gel. The PCR generated cDNAs were cloned into the pCR-TRAP cloning system (GenHunter) using blunt end ligation and expressed in competent cells. Transformed cells were plated on LB plates containing 20 µg/ml tetracycline and incubated overnight at 37°C. The presence of cDNA inserts was screened for by using Tetracycline resistance (TetR) and checked by colony-PCR according to the pCR-TRAP protocol. The use of a phage Lambda repressor gene by this cloning system allows only those plasmids that have a cDNA insert to acquire TetR. A number of insert positive colonies were grown overnight in liquid culture (LB medium + 20 µg/ml tetracycline) and the plasmid DNA harvested the next day using QIAGEN miniprep system. Plasmid DNA was sequenced at the MOBIX facility at McMaster.
Northern hybridization.
The cDNA inserts were excised by Hind III restriction enzyme digestion and separated on a 1.7% agarose gel. The cDNA is recovered from the gel using the QIAEX II gel extraction kit (QIAGEN, Chatsworth, CA) and labelled using a random primed labelling kit (Gibco) and subsequently used as a probe in Northern hybridization. The cDNA probe is verified by Northern blot on both control and PLG treated RNA.
Results
We implemented a strategy for identifying genes involved in the signal transduction pathway of the novel neuro-peptide PLG which employed the differential display of mRNA by RT-PCR technique. An overview of this technique is given in Figure 1.
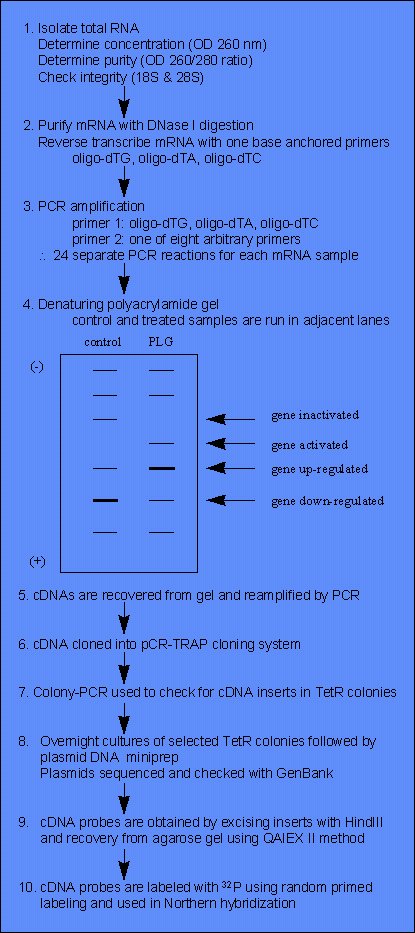 Figure 1. Flow chart of differential display by RT-PCR technique.
Figure 1. Flow chart of differential display by RT-PCR technique.
Approximately 2400 genes were screened using the GeneHunter Kit, and we found three bands which were down-regulated due to treatment with PLG. Figures 2 and 3 are ddPCR gels which are representative of the results we obtained using this technique.
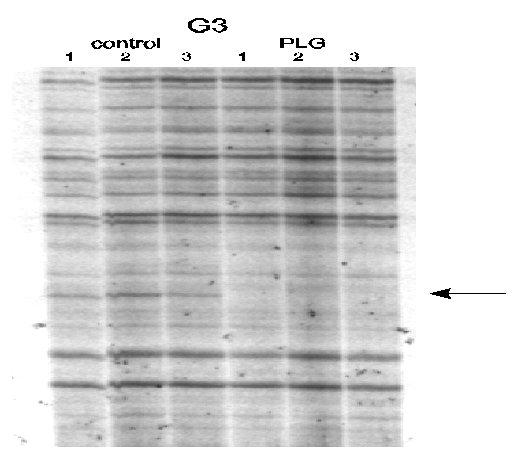 Figure 2. Representative sequencing gel of ddPCR fragments differentially regulated in striatal RNA samples from control and PLG treated rats. It can be seen that the indicated band is absent in the three PLG lanes.
Figure 2. Representative sequencing gel of ddPCR fragments differentially regulated in striatal RNA samples from control and PLG treated rats. It can be seen that the indicated band is absent in the three PLG lanes.
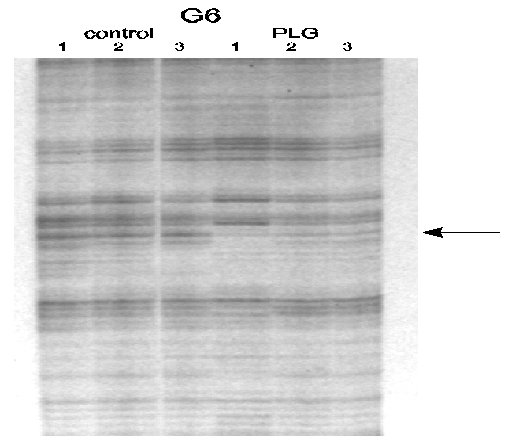 Figure 3. Representative sequencing gel of ddPCR fragments differentially regulated in striatal RNA samples from control and PLG treated rats. It can be seen that the indicated band is absent in the three PLG lanes.
Figure 3. Representative sequencing gel of ddPCR fragments differentially regulated in striatal RNA samples from control and PLG treated rats. It can be seen that the indicated band is absent in the three PLG lanes.
The bands were excised and the cDNAs were recovered and cloned into the pCR-Trap vector. Sequencing of the cDNAs revealed that some of the bands consisted of greater than 1 cDNA species. In total, 6 unique cDNAs were recovered; ranging in size from 133 bp to 228 bp (see table 1). Comparison of the nucleotide sequences we obtained with those in Genbank revealed that three of the cDNAs matched well with known genes (PRG1, PRG3 and PRG6), while three produced poor matches (PRG1, PRG3 and PRG6). The results of the Genbank search are summarized in Table 1.
----------------------------------------------------------------------------
cDNA Length Match Length Blastn score/ NEAREST HOMOLOGY
(bp) (bp) Expect Value Gene sequence match (accession #)
----------------------------------------------------------------------------
PRG1 225 181/187;96% 323/7e-87 Homo sapiens full length insert
cDNA clone ZD50B08 (AF086296)
PRG2 228 43/49;87% 50/1e-4 Genomic sequence from Mouse 11,
33/36;91% complete sequence (AC002121)
43/50;86%
45/54;83%
PRG3 224 168/187;89%* 234/5e-60 Mus musculus mRNA for protein L,
partial cds (AB009392)
PRG4 133 17/17;100% 34/4.8 Caenorhabditis elegans cosmid
T01B7, complete sequence (Z66499)
PRG5 166 20/20;100% 40/0.1 Human Chromosome X, complete
sequence (AC002418)
PRG6 166 88/97;90% 121/3e-26 Mus musculus major histocompatibility
locus class II region (AF050157)
----------------------------------------------------------------------------
TABLE 1. Summary of Genbank search (conducted Oct. 6, 1998) information for PLG regulated gene sequences (PRG1-PRG6) obtained using ddPCR. All of the identified ddPCR fragments were down-regulated as a result of PLG treatment. The table provides the length of the ddPCR fragments obtained as well as the match information (length and percentage of match, score and expect values) for the best match reported by BLSTN 2.0.5. The best matches are given along with their genbank accession numbers. *Indicates match had gaps = 3/187 (1%).
Northern blot analysis is used to confirm the expression of the differentially regulated cDNAs. Figure 4 is a northern blot which confirms the expression of PRG1 as well as comparison to the expression of ß-actin in the same samples.
 Figure 4. Northern hybridization of probes for PRG1 and ß-actin to striatal RNA (20 µg) from control (lanes 1, 2 and 5) and PLG (lanes 2, 4 and 6) treated animals.
Figure 4. Northern hybridization of probes for PRG1 and ß-actin to striatal RNA (20 µg) from control (lanes 1, 2 and 5) and PLG (lanes 2, 4 and 6) treated animals.
Figure 5 shows the expression of PRG1 as a percentage of ß-actin, and confirms that the expression of PRG1 was decreased in the striatum of animals treated with PLG.
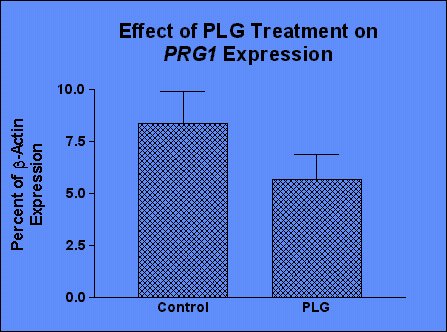 Figure 5. Expression of PRG1 in control and PLG treated rat striatum.
Figure 5. Expression of PRG1 in control and PLG treated rat striatum.
Figure 6 is a northern blot using the cDNA PRG3 as a probe. The probe was hybridized against rat RNA (hypothalamus,hippocampus, pre-frontal cortex and cerebellum) as well as RNA from the human neuroblastoma SH-SY5Y cell line. The probe detected three bands in the rat RNA and two bands in the human RNA. This cDNA is known to match well with the sequences for the heterogeneous nuclear RNP protein L in the mouse and human, so it was expected to hybridize with both rat and human RNA. The human form of protein L RNA is 2033 bp in length while the mouse form is 958 bp in length. It is expected that the rat form of the RNA should be more similar to that of the mouse than of the human. Therefore, the band most likely to be that of protein L is band #3 in the rat and band #2 in the human RNA. Since protein L belongs to a large family of ribonuclear proteins, the other bands detected are likely to be other subtypes of ribonuclear proteins.
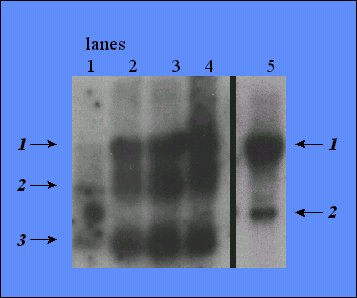 Figure 6. Northern hybridization of probe for PRG3 to RNA from rat hypothalamus (5 µg), hippocampus (17.6 µg), pre-frontal cortex (23.6 µg) and cerebellum (40 µg) (lanes 1, 2, 3 and 4 respectively) and human neuroblastoma cells (SH-SY5Y; 40 µg, lane 5).
Figure 6. Northern hybridization of probe for PRG3 to RNA from rat hypothalamus (5 µg), hippocampus (17.6 µg), pre-frontal cortex (23.6 µg) and cerebellum (40 µg) (lanes 1, 2, 3 and 4 respectively) and human neuroblastoma cells (SH-SY5Y; 40 µg, lane 5).
Additionally, a Norther blot was performed using the cDNA PRG3 as a probe. This cDNA did not hybridize with RNA from rat hypothalamus, hippocampus, pre-frontal cortex cerebellum as well as SH-SY5Y RNA. This indicates that the corresponding RNA is either non-existent or expressed only in the striatum. Further testing will clarify this.
Discussion and Conclusion
We have successfully used ddPCR to identify genes whose expression is regulated by the endogenous neuro-peptide PLG. Although none of the ddPCR fragments we obtained were exact matches with previously identified genes, three of the cDNAs displayed significant homology known gene sequences. This suggests that we have identified either rat homologs of genes cloned in other organisms, or additional members of a family of genes. We are currently in the process of confirming the expression of the cDNAs we have identified, and have obtained positive results for two of the three cDNAs tested to date. Subsequent experiments will involve full-length cloning of those cDNAs confirmed to be expressed in the striatum and regulated by PLG treatment. These experiments will allow us to characterize the role of the novel genes we have obtained herein in mediating the biological effects of PLG. This information will hopefully assist in elucidating the signal transduction mechanisms of PLG.
CONCLUSIONS.
* ddPCR experiments identified 6 unique cDNA species to be down-regulated by PLG treatment
* Sequencing revealed that all cDNAs were previously unidentified in rat
* Three of the cDNAs had poor homology with known genes
==> suggesting novel genes
* Three cDNAs had significant homology with previously identified genes from other species
==> suggesting rat homologs of genes
* Confirmation of expression by Northern hybridization has revealed that two of the three cDNAs tested to date are expressed in rat brain
* Expression of PRG1 has been confirmed to be down-regulated by PLG
References
- Rajakumar G, Naas F, Johnson RL, Chiu S, Yu KL, Mishra RK. Down-regulation of haloperidol-induced striatal dopamine receptor supersensitivity by active analogues of L-prolyl-L-leucyl-glycinamide (PLG). Peptides 1987; 8: 855-861.
- Chiu P, Rajakumar G, Chiu S, Johnson RL, Mishra RK. Mesolimbic and striatal dopamine receptor supersensitivity: Prophylactic and reveral effects of L-prolyl-L-leucyl-glycinamide (PLG). Peptides 1985; 6: 179-183.
- Chiu S, Paulose CS, Mishra RK. Neuroleptic drug-induced dopamine receptor supersensitivity: antagonism by L-prolyl-L-leucyl-glycinamide. Science 1981; 214: 1261-1262.
- Chiu S, Paulose CS, Mishra RK. Effect of L-prolyl-L-leucyl-blycinamide (PLG) on neuroleptic-induced catalepsy and dopamine/neuroleptic receptor binding. Peptides 1981; 2: 105-111.
- Chiu S, Mishra RK. Antagonism of morphine-induced catalepsy by L-prolyl-L-leucyl glycinamide. Eur J Pharmacol 1979; 53: 119-125.
- Ott MC, Mishra RK, Johnson RL. Modulation of dopaminergic neurotransmission in the 6-hydroxydopamine lesioned rotational model by peptidomimetic analogues of L-prolyl-l-leucyl-glycinamide. Brain Res 1996; 737: 287-291.
- Baures PW, Ojala WH, Gleason WB, Mishra RK, Johnson RL. Design, synthesis, X-ray analysis, and dopamine receptor-modulating activity of mimics of the "C5" hydrogen-bonded conformation in the peptidomimetic 2-oxo-3-(R)-[2(S)-pyrrolidinylcarbonyl)amino]-1- pyrrolidineacetamide. J Med Chem 1994; 37: 3677-3683.
- Liang P, Pardee AB. Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science 1992; 257: 967-971.
- Nair RMG, Kastin AJ, Schally AV. Isolation and structure of hypothalamic MSH release-inhibbiting hormone. Biochem Biophys Res Comm 1971; 43: 1376-1381.
- Celis ME, Taleisnik S, Walter R. Regulation of formation and proposed structure of the factor inhibiting the release of alpha-MSH. Proc Natl Acad Sci 1971; 68: 1428-1433.
- Srivastava LK, Bajwa SB, Johnson RL, Mishra RK. Interaction of L-prolyl-L-leucyl glycinamide with dopamine D2 receptor: Evidence for modulation of agonist affinity states in bovine striatal membranes. J Neurochem 1988; 50: 960-968.
- Plotnikoff NP, Kastin AJ, Anderson MS, Schally AV. DOPA potentiation by MIF. Life Sci 1971; 10: 1279-1283.
- Barbeau A. Potentiation of levoodopa effect by intravenous L-prolyl-L-leucyl-glycine amide in man. Lancet 1975; ii: 683-684.
- Barbeau A, Roy M, Gonce M, Labreoque L. Newer therapeutic approaches in Parkinson's Disease. In: Poirier LJ, Sourkes TL, Bedard PJ, eds.Advances in Neurology. New York: Raven Press, 1978; 433-437.
- Ehrensing RH. Lithium and M.R.I.H. in tardive dyskinesia. Lancet 1974; 2: 1459-1460.
- Ehrensing RH, Kastin AJ, Larsons PF, Bishop GA. Melanocyte stimulating hormone release inhibiting factor I and tardive dyskinesia. Dis Nerv Syst 1977; 38: 303-307.
- Hara C, Kastin AJ. Biphasic effects of MIF-1 and Tyr-MIF-1 on apomorphine-induced stereotypy in rats. Pharmacology, Biochemistry & Behavior 1986; 25: 757-761.
- Smith JR, Morgan M. The effects of prolyl-leucyl-glycine amide on drug-induced rotation in lesioned rats. General Pharmacology 1982; 13: 203-207.
- Kostrzewa RM, Kastin AJ, Sobrain SK. Potentiation of apomorphine action in rats by l-prolyl-l-leucyl-glycine amide. Pharmacol Biochem Behav 1978; 9: 375-378.
- Plotnikoff NP, Kastin AJ. Pharmaclogical studies with a tripeptide L-prolyl-L-leucyl-glycinamide. Arch Int Pharmacodyn Ther 1974; 211: 211-224.
- Plotnikoff NP, Kastin AJ. Oxotremorine antagonism by prolyl-leucyl-glycine amide administered by different routes and with several anticholinergics. Pharmac Biochem Behav 1974; 2: 417-419.
- Huidobro-Toro JP, Scotti de Carolis A, Longo VG. Actions of two hypoothalamic factors (TRH, MIF) and of angiotensin II on behavioral effects of L-DOPA and 5-hydroxytryptophan in mice. Pharmac Biochem Behav 1974; 2: 105-109.
- Bjorkman S, Sievertsson H. On the optimal dosage of Pro-Leu-Gly-NH2 (MIF) in neurological test and clinical use. Naunyn Schmiedeberg's Arch Pharmacol 1977; 298: 79-81.
- Sheng JG, Xu DL, Yu HZ, Xu XR, Tang QM. Partial protection from the dopaminergic neurotoxin 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) by pro-leu-gly-NH2 (PLG; MIF-1). Life Sci 1987; 40: 2007-2010.
- Yu K-L, Rajakumar G, Srivastava LK, Mishra RK, Johnson RL. Dopamine receptor modulation by conformationally constrained analogues of pro-leu-gly-NH2. J Med Chem 1988; 31: 1430-1436.
- Subasinghe NL, Bontems RJ, McIntee E, Mishra RK, Johnson RL. Bicyclic thiazolidine lactam peptidomimetics of the dopamine receptor modulating peptide Pro-Leu-Gly-NH2. Journal of Medicinal Chemistry 1993; 36: 2356-2361.
- Johnson RL, Rajakumar G, Mishra RK. Dopamine receptor modulation by Pro-Leu-Gly-NH2 analogues possessing cyclic amino acid residues at the C-terminal position. J Med Chem 1986; 29: 2100-2104.
- Johnson RL, Rajakumar G, Yu K-L, Mishra RK. Synthesis of Pro-Leu-Gly-NH2 analogues modified at the prolyl residue and evaluation of their effects on the receptor binding activity of the central dopamine receptor agonist, ADTN. J Med Chem 1986; 29: 2104-2107.
- Johnson RL, Botems RJ, Yang KE, Mishra RK. Synthesis and biological evaluation of analogues of Pro-Leu-Gly-NH2 modified at the leucyl residue. J Med Chem 1990; 33: 1828-1832.
- Genin MJ, Mishra RK, Johnson RL. Dopamine receptor modulation by a highly rigid spiro bicyclic peptidomimetic of Pro-Leu-Gly-NH2. J Med Chem 1993; 36: 3481-3483.
- Sreenivasan U, Mishra RK, Johnson RL. Synthesis and dopamine receptor modulating activity of lactam conformationally constrained analogues of Pro-Leu-Gly-NH2. J Med Chem 1993; 36: 256-263.
- Mishra RK, Srivastava LK, Johnson RL. Modulation of high-affinity CNS dopamine D2 receptor by L-pro-L-leu-glycinamide (PLG) analogue 3(R)-(N-L-prolylamino)-2-oxo-1- pyrrolidineacetamide. Prog Neuro-Psychopharmacol Biol Psychiat 1990; 14: 821-827.
- Kahn RS, Davis KL. New developments in dopamine and schizophrenia. In: Bloom FE, Kupfer DJ, eds.Psychopharmacology: The Fourth Generation of Progress. New York: Raven Press, Ltd. 1995; 1193-1203.
- Burt, D.R., Creese, I., and Snyder, S.H. Antischizophrenic drugs: Chronic treatment elevates dopamine receptor binding in brain. Science 196:326-328, 1977.
- Ott, M.C., Johnson, R.L., and Mishra, R.K. PLG and a peptidomimetic of PLG antagonize haloperidol induced c-fos expression within the striatum and nucleus accumbens. J. Neurochem. (submitted) 1997.
| Discussion Board | Previous Page | Your Poster Session |