Neuropharmacology Poster Session
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
An understanding of the mechanisms involved in the development of sensitisation and tolerance to dopaminergic drugs is an area of growing interest for drug therapy in Parkinson's disease, as well as for animal models of schizophrenia and addiction. The psychomotor stimulant, (+)-4-propyl-9-hydroxynaphthoxazine (PHNO) is a potent dopamine (DA) agonist with a distinct selectivity for the D2 receptor subtype (Martin et al. 1984; Seeman et al. 1993). When PHNO is continuously administered to rats, the development of both tolerance and sensitisation occurs, dependent upon the light-dark cycle (Martin-Iverson et al. 1988a; Martin-Iverson et al. 1988b). Rats given a continuous (24-hour) infusion of PHNO (0.005 mg/h) exhibit tolerance to the behavioural effects of PHNO during the light period of a 12 hr light-dark cycle, and show sensitisation to the effects of PHNO during the dark period (Martin-Iverson et al. 1988a; 1988b). By reversing the light-dark schedule (from lights on from 09:00-21:00 to lights on from 21:00-09:00) tolerance to the effects of PHNO shifts (within 3-4 days) to follow 'daylight' hours and sensitisation switches to the opposite 'night-time' schedule (lights on from 09:00-21:00 rather than 21:00 to 09:00) (Martin-Iverson et al. 1988a).
Sensitisation and tolerance are not solely dependent on light cues as rats maintained in constant darkness exhibit sensitisation during active periods (at the peak of the rhythm) and tolerance during the resting phases or rhythm nadir (Martin-Iverson & Yamada 1992). Rats kept under constant light exhibit a disruption of circadian rhythms, and exhibit no motor stimulant effects to PHNO, although peak levels of activity are comparable in magnitude to those rats exhibiting circadian rhythms. As such, the development of sensitisation and tolerance is a function of free-running circadian rhythms in motor activity, rather than the lighting schedule itself, or rate-dependent effects.
Since constant light atrophies the pineal gland that secretes melatonin (producing a virtual pinealectomy) and blocks motor stimulant effects of PHNO, it is possible that daytime tolerance to PHNO in rats under 12:12 h lighting conditions is due to a reduction in melatonin secretion, and nocturnal sensitisation may be due to increased melatonin secretion. It is hypothesised that the pineal hormone melatonin may mediate responsiveness to the variations in the motor stimulant effects of continuously administered PHNO.
A recent study has shown that reversal of diurnal tolerance to continuous PHNO infusion can be achieved by injecting rats with 2-iodo-melatonin, a potent melatonin receptor agonist (Munro 1997). This evidence supports the possibility that inhibition of melatonin release from the pineal gland is responsible for diurnal tolerance and increasing melatonin release responsible for nocturnal sensitisation. As such it is predicted that rats without a pineal gland and hence no melatonin, will not develop sensitisation to continuous PHNO infusion.
In the present study, pinealectomies were performed to assess the hypothesised role of nocturnal melatonin release in the development of sensitisation to continuous PHNO administration. It is predicted that tolerance to continuous infusion of PHNO will develop in sham-operated rats during the light hours and sensitisation will develop during the dark hours, as assessed by automated measures of rearing behaviour. It is also predicted that pinealectomy will block nocturnal sensitisation in animals receiving a continuous infusion of PHNO and if so, administration of 2-iodo-melatonin should reinstate sensitisation in the pinealectomised animals.
Materials and Methods
46 Experimentally na�ve, Sprague-Dawley rats weighing 371 - 523 g were housed in cages on a timer controlled 12:12 h light/dark cycle (lights on 18:30-06:30). Dark illumination was provided by a red incandescent light (1 lux) during nocturnal injection procedures and light illumination by an overhead full-spectrum fluorescent light (160 lux). Temperature was maintained at 22+-2�C. The animals had unlimited access to food (Glen Forrest Stockfeeders) and water. The animals were individually housed in motor activity testing cages. The cage dimensions were 21 (W) by 18(H) by 33(L) cm.. Each individual cage was positioned between four infra-red photocell beams placed near the top of the cages, which provided continuous monitoring of rears. A computer recorded nonsuccessive photocell interruptions cumulated over 1-hour blocks (rears).
(+)-4-propyl-9 hydroxynaphthoxazine hydrochloride (PHNO, Merck, Sharp and Dohme) was dissolved in distilled water and ALZET osmotic pumps (Model #2002) were filled with the solution providing a release rate of approximately 0.005 mg/hr. 2-Iodo-melatonin was dissolved in 7.25% Tween 80 for a dose of 0.5 mg/kg. All drug dosages are expressed in weights of their salts.
The animals were habituated to cages for 2 weeks prior to surgery. In rats anaesthetised with Nembutol (maintained with halothane), pinealectomies were performed following a modification of the method of Pohlmeyer, Reuss and Baum (1994). All procedures were followed in the sham-operated controls except that the pineal gland was left intact. Osmotic pumps were inserted S.C. in the midscapular regions immediately after the pinealectomies while the rats remained anaesthetised.
The groups in this experiment were Sham surgery + Vehicle (S+V; n=11), Sham surgery + PHNO (S+P; n=12), Pinealectomy + Vehicle (X+V; n=11) and Pinealectomy + PHNO (X+P; n=12). The night of day 9 marked the beginning of the second phase of the experiment. At 20:30 h on day 9 for each animal, an IP injection of either 2-iodo-melatonin or vehicle was administered to each rat. On the following night, at 20:30 h they received a vehicle injection if they had received the drug the night before or they received the drug injection if they had previously received the vehicle injection. Thus, each animal was injected with both the vehicle solution and 2-iodo-melatonin solution, with the order of injections counter-balanced across animals.
The dependent variables for the first analysis were day 1, day 7, night 2 and night 8 were mean hourly rears as a percentage of baseline hourly rates to equate night and day measures and to assess possible effects of the surgeries relative to presurgery baseline. A mixed-model ANOVA was used with two between factors, each with two levels: surgery (pinealectomy or sham) and drug (PHNO or vehicle), and two within factors in the analysis (2 levels each), which were Light Condition (day or night) and treatment day (two levels: 1 and 7 for day; 2 and 8 for night).
Rearing counts from the hour before the injection procedure and one hour after the procedure were analysed to examine the effect of the 2-iodo-melatonin injections. The ANOVA had two between factors each with two levels: Surgery (sham or pinealectomy), Infused Drug (vehicle or PHNO), and two within factors each with two levels, Injected Drug (vehicle or 2-iodo-melatonin) and Treatment Time (pre or post Injection). Specific pairwise comparisons were made using critical differences derived from the Multiple F-test (Kiess 1989).
Results
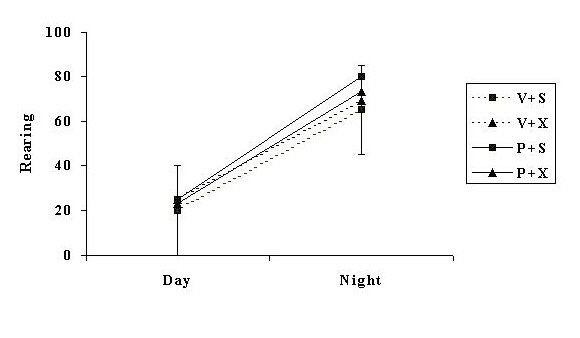
Autopsy revealed that all but three of the attempted pinealectomies successfully removed the pineal gland. These three animals were placed in the sham group for the analyses. There were no significant differences in the level of activity between groups before surgery (Figure 1).
 Click to enlarge
Fig. 1: Mean rears averaged over the two weeks prior to surgery (baseline measure), for groups of rats subsequently treated with infusions of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X). The error bar represents the critical difference between means (p < 0.05).
Click to enlarge
Fig. 1: Mean rears averaged over the two weeks prior to surgery (baseline measure), for groups of rats subsequently treated with infusions of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X). The error bar represents the critical difference between means (p < 0.05).
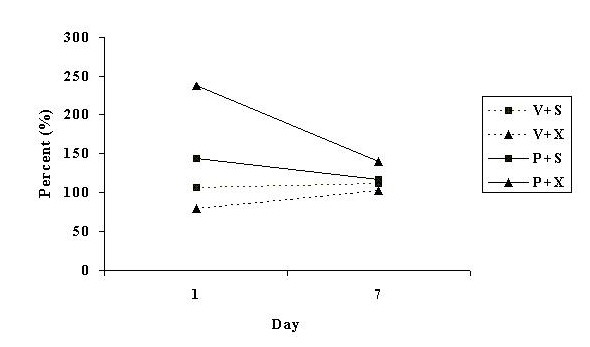
Mean rears as a percent of baseline during the light phase of day 1 and day 7 of treatment, for groups of rats given a sham or pinealectomy operation and treated with continuous infusions of vehicle or PHNO are illustrated in Figure 2.
 Click to enlarge
Fig. 2: Diurnal rears as a percent of its baseline measure, for day 1 and day 7, for groups of rats treated with infusions of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X). PHNO treated animals exhibit a decrease in rears from day 1 to day 7 (tolerance).
Click to enlarge
Fig. 2: Diurnal rears as a percent of its baseline measure, for day 1 and day 7, for groups of rats treated with infusions of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X). PHNO treated animals exhibit a decrease in rears from day 1 to day 7 (tolerance).
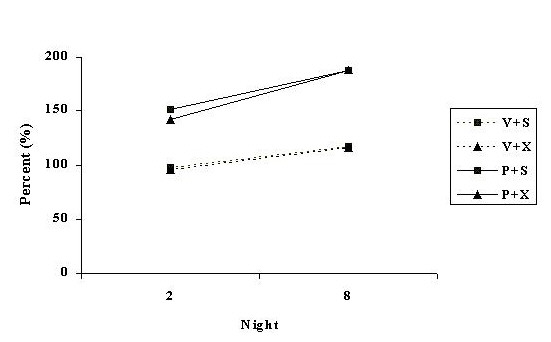
The corresponding data for night 2 and night 8 are depicted in Figure 5.
 Click to enlarge
Fig. 3: Nocturnal rears expressed as a percent of its baseline for nights 2 and 8, in groups of rats treated with continuous infusion of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X).
Click to enlarge
Fig. 3: Nocturnal rears expressed as a percent of its baseline for nights 2 and 8, in groups of rats treated with continuous infusion of vehicle and given a sham operation (V + S), infused with vehicle and pinealectomised (V + X), infused with PHNO and given a sham operation (P + S), or infused with PHNO and pinealectomised (P + X).
ANOVA revealed a significant surgery x drug x light condition interaction [F(1,42) = 4.08, p < .05]. Figures 2 and 3 indicate that this interaction is due to the observation that PHNO was more effective at increasing rears on day 1 than in the sham-operated controls or in the lesioned animals at night. ANOVA also revealed a significant drug x light condition x day interaction [F(1, 42) = 6.90, p < .025]. As Figs. 2 and 3 illustrate, this interaction is due to the PHNO treated animals, as compared to the sham animals, to have more rears, on day 1 compared to day 7 but more rears on night 8 compared to night 2. The decrease in the behavioural response from day 1 to day 7 is characteristic of tolerance, and the increase from night 2 to night 8 is sensitisation. The four-way interaction of drug x surgery x light condition x treatment day was not significant [F(1, 42) = 1.22, p > .2]. This interaction was expected to reach significance if melatonin had been involved in the development of tolerance or sensitisation to the effects of PHNO.
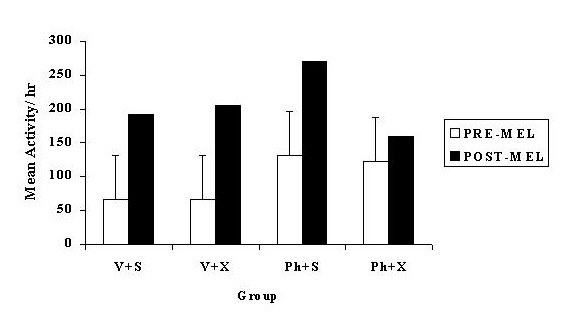
Figure 4 shows the effects of the 2-iodo-melatonin injections on nocturnal rears. Rats generally increased rearing after the injection as compared to before, indicated by a significant main effect of pre/post-injection [F(1, 42) = 46.87, p < .001]. This effect is illustrated in Figure 4, where planned comparisons indicate that all groups significantly increased their activity post injection except the
 Click to enlarge
Fig. 4: Mean nocturnal rears 1 hour before (PRE-MEL) and 1 hour after (POST-MEL) 2-iodo-melatonin injections given to pinealectomised or sham operated rats, administered continuous PHNO (0.005 mg/h) or vehicle through a mini-pump. The treatment groups are: Vehicle + Sham (V + S), Vehicle + Pinealectomy (V + X), PHNO + Sham (P + S), PHNO + Pinealectomy (P + X). The error bars represent the critical difference between means (p<0.05).
Click to enlarge
Fig. 4: Mean nocturnal rears 1 hour before (PRE-MEL) and 1 hour after (POST-MEL) 2-iodo-melatonin injections given to pinealectomised or sham operated rats, administered continuous PHNO (0.005 mg/h) or vehicle through a mini-pump. The treatment groups are: Vehicle + Sham (V + S), Vehicle + Pinealectomy (V + X), PHNO + Sham (P + S), PHNO + Pinealectomy (P + X). The error bars represent the critical difference between means (p<0.05).
pinealectomised animals receiving PHNO. However, neither the main effect of pinealectomy nor any interaction involving pinealectomy was significant. The pre/post-injection analysis of the vehicle injections produced a different pattern of results as compared to the effect of the 2-iodo-melatonin injections. The pre/post-injection analysis was not significant, indicating that activity did not significantly differ post vehicle injection as compared to the hour before the injection. However, ANOVA did reveal a significant surgery x pre/post-injection interaction [F(1, 42) = 4.38, p < .05]. As illustrated in Figure 5, planned comparisons indicate that the activity of both sham groups was significantly increased after the injection procedure as compared to the hour before the injection (p < .05). In contrast, planned comparisons showed that the activity of the pinealectomised groups did not significantly differ between the hour before and after the injection (p < .05). The pinealectomised group receiving PHNO actually exhibited a non-significant decrease in their activity level post injection.
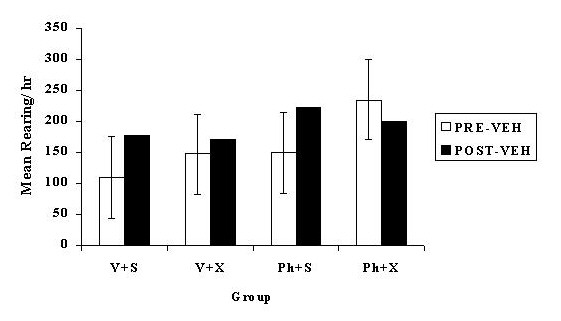 Click to enlarge
Fig. 5: Mean nocturnal rears 1 hour before (PRE-VEH) and 1 hour after (POST-VEH) vehicle injections given to pinealectomised or sham operated rats administered continuous PHNO (0.005 mg/h) or vehicle via osmotic pump. The error bars represent the critical difference. The groups are V + S: Vehicle pump + Sham operation, V + X: Vehicle pump + pinealectomy, P + S: PHNO + Sham operation, P + P: PHNO + Pinealectomy. The error bars represent the critical difference between means (p<0.05).
Click to enlarge
Fig. 5: Mean nocturnal rears 1 hour before (PRE-VEH) and 1 hour after (POST-VEH) vehicle injections given to pinealectomised or sham operated rats administered continuous PHNO (0.005 mg/h) or vehicle via osmotic pump. The error bars represent the critical difference. The groups are V + S: Vehicle pump + Sham operation, V + X: Vehicle pump + pinealectomy, P + S: PHNO + Sham operation, P + P: PHNO + Pinealectomy. The error bars represent the critical difference between means (p<0.05).
Discussion and Conclusion
This study shows that melatonin release is not involved in the mediation of diurnal tolerance or nocturnal sensitisation to the behavioural effects of the selective DA D2 agonist PHNO. Rats were given either a sham or pinealectomy operation, and implanted with pumps which infused either vehicle (distilled water) or PHNO (0.005 mg/h). It was hypothesised that if endogenous melatonin were responsible for the behavioural effects of PHNO, then pinealectomised animals would not develop nocturnal sensitisation to its effects. In contrast to this prediction, the current results indicate that the motor stimulant effects of PHNO are not influenced by a lack of melatonin resulting from pinealectomy, as measured by the automated counting of rears. Indeed, the only effect of pinealectomy observed from day 1 to day 7 or night 2 to night 8 of drug treatment was a daytime increase in rears on day 1. If the pinealectomised animals receiving PHNO had not exhibited the nocturnal sensitisation to PHNO, this would have confirmed the prediction that melatonin in some way mediates the development of behavioural effects to PHNO. As pinealectomised rats developed sensitisation to continuous PHNO infusion, it can be concluded that melatonin is not necessary for nocturnal sensitisation to the behavioural effects of PHNO.
The present results extend previous findings that continuous infusion of PHNO in rats results in the development of tolerance to its locomotor effects during the day and sensitisation to its effects at night (Martin-Iverson et al. 1988a, 1988b) to similar effects on rearing behaviour. The same animals that developed tolerance to the effects of PHNO during the light hours produced an augmentation of the behavioural response (sensitisation) to PHNO's rearing effects in the dark hours. Rearing for the animals receiving PHNO show a significant decrease from day 1 to day 7 (tolerance) and exhibit an increase from night 2 to night 8 (sensitisation).
The current results question previous conclusions pertaining to the function of melatonin in mediating the behavioural effects produced by continuous infusion of PHNO. Munro (1997) concluded that diurnal tolerance and nocturnal sensitisation to PHNO are mediated to some degree by melatonin, after finding that diurnal tolerance to PHNO can be reversed by injecting animals with 2-iodo-melatonin. The present data suggest that the injection procedure is a stressful event, and as stress has been shown to reverse tolerance to the effects of PHNO (Martin-Iverson et al. 1988a), the influence of melatonin and stress were most likely confounded in Munro's study (1997). The present results indicate that the findings of Munro were likely the augmentation of stress effects, rather than the reversal of tolerance to PHNO.
Indeed, the main effect of pinealectomy observed in the present study was a reduction in the stress response to vehicle injection. This response was significantly re-instated by 2-iodo-melatonin injections in vehicle treated rats. Furthermore, a trend towards a decrease in activity after vehicle injections in PHNO infused rats was reversed to a trend towards an increase by 2-iodo-melatonin. There is independent evidence which supports a role for melatonin in stress, as studies have shown that melatonin levels have been significantly elevated after exercise-induced stress in women (Carr et al. 1981), and after swim-induced stress in rats (Tenn and Niles 1993).
As it was found that melatonin is probably not involved in the development of the circadian patterns of tolerance and sensitisation, which occur in response to continuous administration of PHNO, the question remains as to what does mediate the development of these drug responses. Research has shown that the expression of PHNO's behavioural effects is a function of a synergistic interaction between endogenous dopamine and exogenous receptor agonists acting on both D1 and D2 dopamine receptors (Martin-Iverson et al. 1988b; Martin-Iverson & Yamada 1992; Yamada & Martin-Iverson 1991). The evidence for this hypothesis is indicated by the finding that PHNO's motor stimulant effects are blocked by co-administration of D1 receptor antagonist, SCH 233390 (Martin-Iverson et al. 1988b). In addition, it has been shown that co-administration of the D1 receptor agonist SKF 38393, reverses tolerance to the effects of PHNO (Martin-Iverson et al. 1988b). This evidence suggests that daytime tolerance to PHNO results from a decrease in endogenous dopamine release such that activation of D1 receptors is reduced, but at night the activation of D1 receptors is increased, producing sensitisation. The present data indicate that whatever regulates the responsiveness of dopamine release regulating daytime tolerance and nocturnal sensitisation to PHNO, it is not melatonin. However, melatonin does appear to play a role in stress effects on rearing behaviour.
References
- Carr DB, Reppert SM, Bullen B, Skrinar G, Beitins I, Arnold M, Rosenblatt M, Martin JB, McWarren JW (1981) Plasma melatonin increases during exercise in women. Journal of Clinical Endocrinology, 10: 126-129.
- Kiess HO (1989) Statistical concepts for the behavioural sciences. Allyn and Bacon, Toronto.
- Martin GE, Williams M, Pettibone DJ, Yarbrough GG, Clineschmidt BV, Jones JH (1984) Pharmacologic profile of a novel potent direct-acting dopamine agonist, (+)-4-propyl-9-hydroxynaphthoxazine [(+)-PHNO]. Journal of Pharmacology and Experimental Theraputics, 230: 569-576.
- Martin-Iverson MT, Iverson SD, Stahl SM (1988b) Long-term motor stimulant effects of (+)-4-propyl-9-hydroxynaphthoxazine (PHNO) a dopamine D-2 receptor agonist: interactions with a dopamine D-1 receptor antagonist and agonist. European Journal of Pharmacology, 149: 25-31.
- Martin-Iverson MT, Stahl SM, Iverson SD (1988a) Chronic administration of a selective dopamine D-2 agonist: factors determining behavioral tolerance and sensitization. Psychopharmacology, 95: 534-539.
- Martin-Iverson MT, Yamada N (1992) Synergistic behavioural effects of dopamine D1 and D2 agonists are determined by circadian rhythms. European Journal of Pharmacology, 215: 119-125.
- Munro J (1997) The impact of circadian melatonin release to the development of tolerance and sensitisation to the dopamine D2 receptor agonist, (+)-4-propyl-9-hydroxynaphthoxazine (PHNO). Honours Thesis University of Western Australia.
- Pohlemeyer, Reuss & Braun (1994) Pinealectomy in the rat. Journal of Experimental Animal Science, 36: 84-88.
- Seeman P, Ulpian C, Larsen RD, Anderson PS (1993) Dopamine receptors labeled by PHNO. Synapse, 14: 254-262.
- Tenn C, Niles LP (1993) Physiological regulation of melatonin receptors in rat suprachiasmatic nuclei: diurnal rhythmicity and effects of stress. Molecular and Cellular Endocrinology, 98: 43-48.
- Yamada N, Martin-Iverson MT (1991) Selective dopamine D1 and D2 agonists independently affect different components of the free-running circadian rhythm of locomotor activity in rats. Brain Research, 538: 310-312.
| Discussion Board | Previous Page | Your Poster Session |