Invited Symposium: Genital Sensation: CNS Targets and Functions in Females
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Introduction
The lordotic response of female mammals displayed during mating is principally triggered by the mechanical stimulation that the male provides to specific skin areas, perineum, vagina and cervix. Estrous cats (Bard, 1935; Diakow, 1971) and rats (Komisaruk, 1978) are suitable models to study reflex responses after appropriate stimulation which mimics stimulation by the male. Thus, the present report summarizes studies related to some somato-somato and somato-visceral reflexes induced by genital stimulation in cat and rat females under different hormonal conditions (intact, ovariectomized, and ovariectomized plus estrogen and progesterone treatment). Hoping to provide that postural and visceral spinal reflexes of females involved in reproduction are modulated by gonadal hormones which action is ejerced at higher nervous level as well as at low spinal levels.
Materials and Methods
Studies in cat: Were done in acute spinal-transected (C1-C2 level) females which previously were maintained under different hormonal conditions: intact (n=6), ovariectomized (Ovx, two weeks, n=9), and Ovx plus estrogen treatment (8-18 days, n=7). 10 g of estradiol benzoate were injected daily. The spinal cord was transected under ether anesthesia. After the lesion, artificial respiration was installed and anesthesia discontinued. Monosynaptic L7 ventral root responses were recorded and analyzed after electrical stimulation to the ipsilateral medial gastrocnemius nerve, tapping to the perivulvar skin, neckgrip manual pressure, and vaginal or cervix stimulation with a glass probe. Bipolar silver-chlorurated electrodes were used for stimulation and recording. The size of the monosynaptic response, which was raised until a polysynaptic response appeared in the recording, was compared before and during the above mentioned stimuli. The percentage increase or reduction was calculated. An analysis of variance and post-hoc Tukey test were carried out for statistical comparion of data.
Studies in rat: Were done in two experiments. Experiment A. Analysis of electromiographyc (EMG) activity of two muscles of the pelvic region, the iliococcygeus and pubococcygeus, and two muscles involved in postural hindleg activity, the psoas major and iliacus. The EMG analysis was performed during mechanostimulation of the perineal skin, clitoral sheat, lower vagina and cervix of virgin Sprague-Dawleys (259-300 g), anesthetized with urethane (1.6 g/kg, ip). Pairs of stainless steel wires (0.1 mm dia) were used for the EMG recordings. Our analysis was based only on the presence-absence of reflex activity. Experiment B. Analysis of the reflexive dilation of the external vaginal orifice (VOD). It was done in virgin Wistar rats (250-300 g) during two tests performed in different stages of their estrous cycle (n=10), or after being ovariectomized for two weeks (Ovx, n=11), or after Ovx and consecutive sc injections of estradiol benzoate (10 g per day for 5 days, n=10), or after secuential sc injection of estradiol benzoate (4 days) and a dose of progesterone (2 mg) administered 4 h before the observation test (n=9). Tapping of the perigenital area in groups of 10 taps was used as the experimental stimulus to induce VODs. The two tests were done two hours after the light was off in the animal room, that had an inverted 12:12 LD cycle. Each test consisted of 20 trials during which the animal was holding by the dorsum to expose the perineal region. During each trial the presence or absence of VOD was recorded. Presence was scored when the external vaginal orifice had more that 1 mm dia aperture. The first test was just to record "spontaneous" VOD’s. The second test was to record VODs provoked by tapping. Data is expressed as percentage and the Fisher Exact Test was done for comparisons.
Results
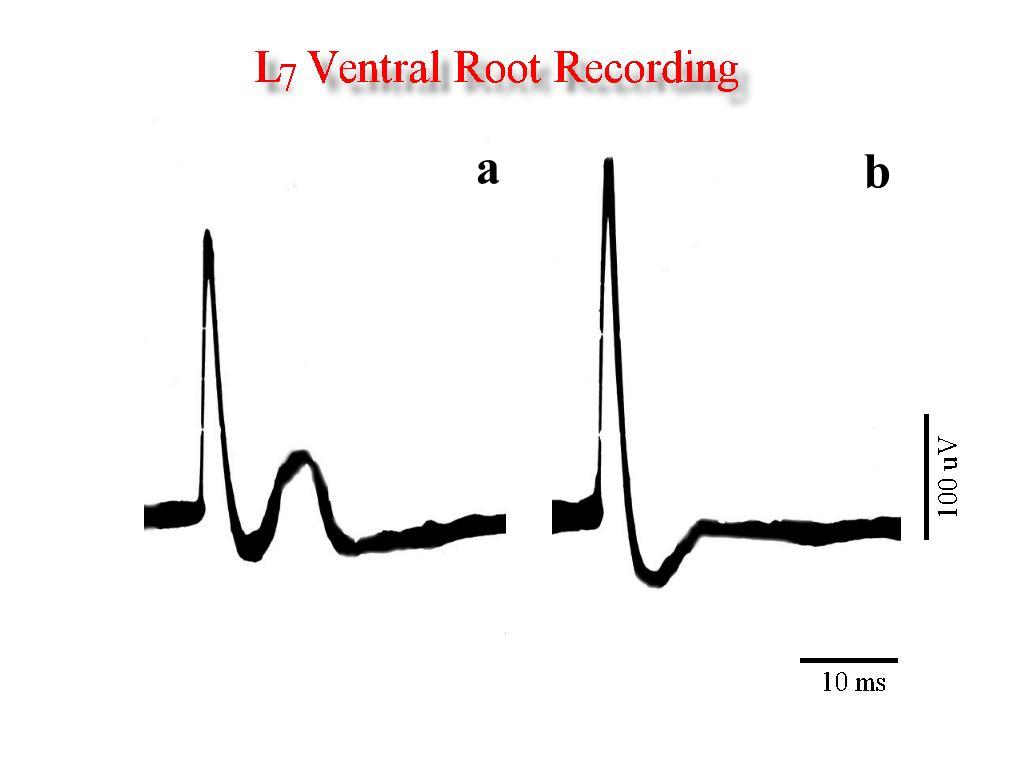
Studies in cats Fig 1a shows typical L7 ventral root responses evoked by electrical stimulation of the ipsilateral left medial gastrocnemius nerve. These responses show the polysynaptic component which was considered to standardize the intensity (thershold) in all animals studied. In 1b it is shown the effect produced during the additional tapping of the perivulvar skin, that is, facilitation of the monosynaptic response and inhibition of the polysinaptic component.
 Click to enlarge
Fig. 1: Ventral root recording to medial gastrocnemius electrical stimulation. Note that tapping of the perivulvar skin area produces facilitation of the monosynaptic response and inhibition of the polysinaptic component.
Click to enlarge
Fig. 1: Ventral root recording to medial gastrocnemius electrical stimulation. Note that tapping of the perivulvar skin area produces facilitation of the monosynaptic response and inhibition of the polysinaptic component.
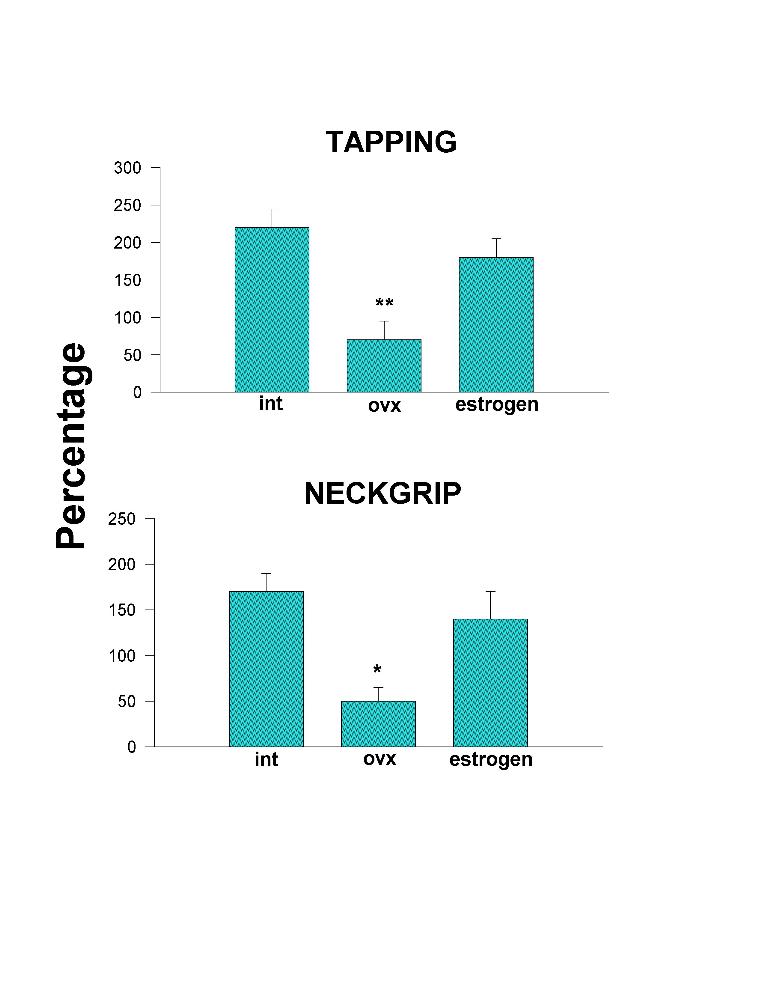
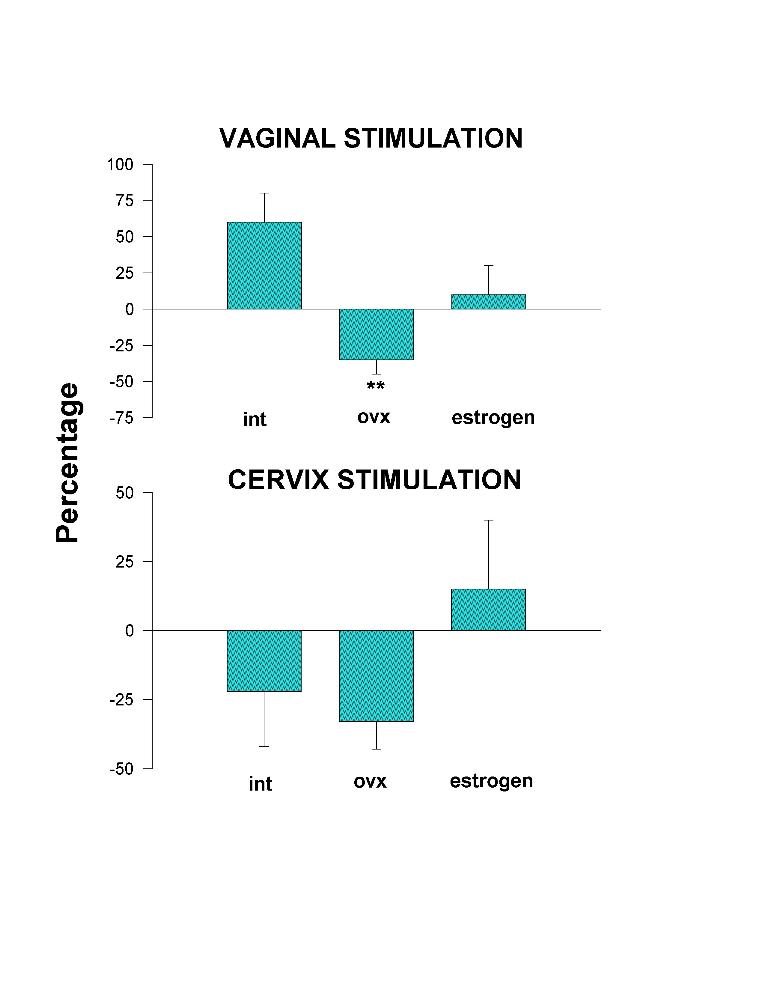
Figs. 2 and 3 show that Ovx reduces significantly the facilitatory effect of tapping and neckgrip stimulation, and inverts to an inhibitory effect the vaginal stimulation. The Ovx also maintains, and even with less variation, the inhibitory effect produced by the cervix stimulation. Administration of estrogen to Ovx cats brings back to intact values the facilitatory effect of tapping, neckgrip, and vaginal stimulation. Outstanding was the estrogen effect upon the cervix stimulation which resulted in facilitation instead of inhibition obtained in either intact or Ovx animals (Fig. 3).
 Click to enlarge
Fig. 2
Click to enlarge
Fig. 2
 Click to enlarge
Fig. 3
Click to enlarge
Fig. 3
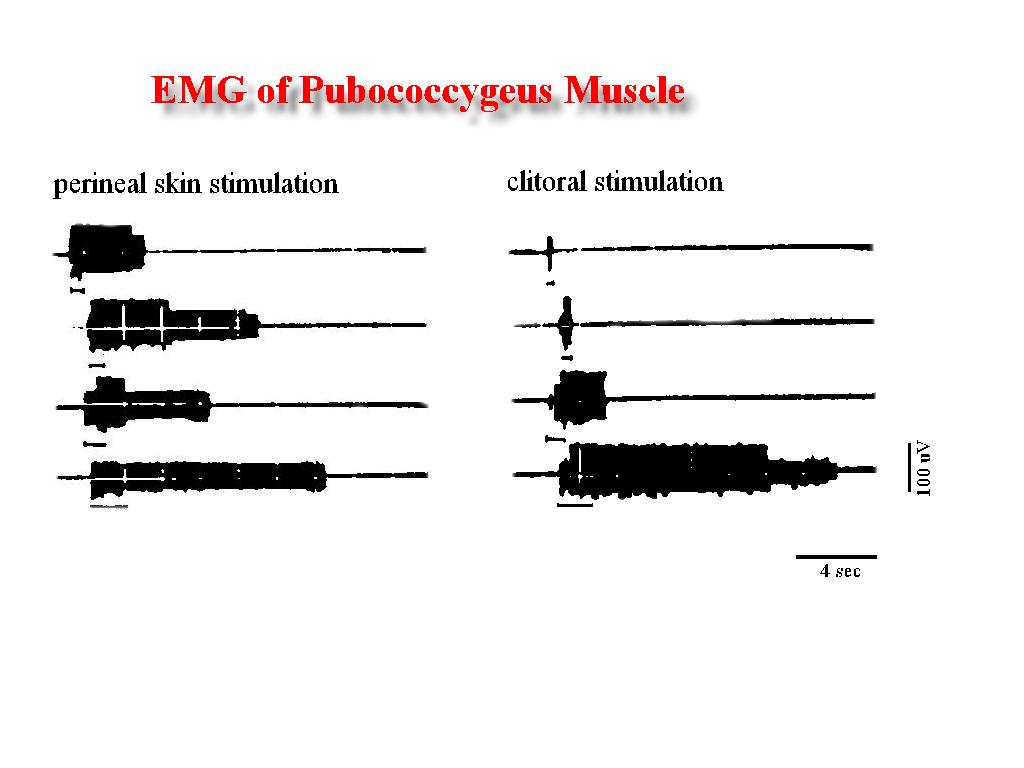
Studies in rats Experiment A. Even that the female rat was anesthetized, the mechanostimulation of the perineal skin, clitoral sheat or lower vagina elicits reflexive EMG activity of the iliococcygeus and pubococcygeus muscles. Distal vagina stimulation by pressuring with a glass probe produce EMG activity that was recorded only during stimulation. However, persisting beyond the cessation of the perineal skin or clitoral sheat traction (afterdischarge) was the characteristic of the EMG activity recorded and was related to the stimulus duration (Fig. 4). Whereas the above stimuli had no observable effect upon the psoas major and iliacus muscles, light touching of the cervix with the intravaginal glass probe produce a strong EMG activity of these two muscles. The activity persist if the light pressure to the cervix persist. However, when this stimulation was secuentially repeated an afterdischarge was recorded (Fig. 5).
 Click to enlarge
Fig. 4
Click to enlarge
Fig. 4
 Click to enlarge
Fig. 5
Click to enlarge
Fig. 5
If during traction of either clitoris or perineal skin the intravaginal glass probe touch the cervix, the EMG activity recorded in the ilio- and pubococcygeus muscles is immediately inhibited while the nule basal EMG activity of the iliacus and psoas major muscles is transformed to a strong EMG activity (Fig. 6).
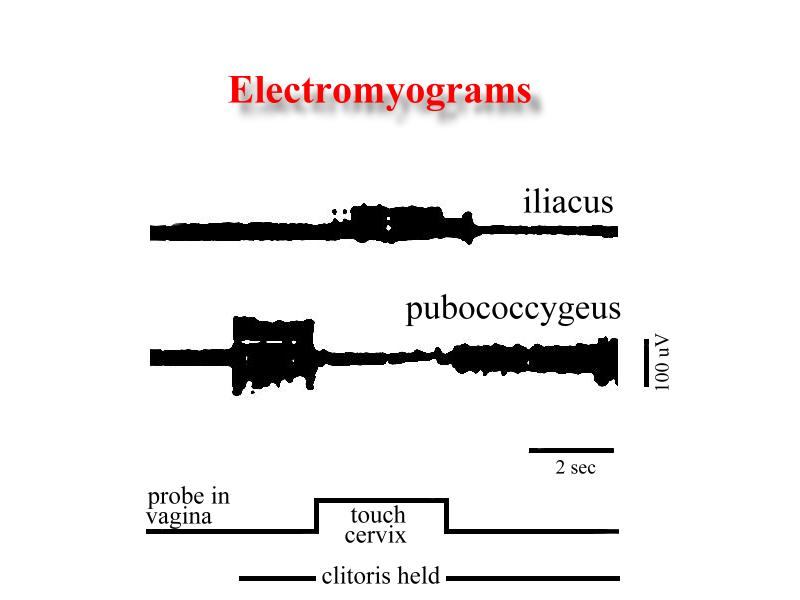 Click to enlarge
Fig. 6
Click to enlarge
Fig. 6
The possible sensory pathways related with this reflexive EMG activity was explored doing nerve sections. Thus, bilateral transection of the viscerocutaneous branch of the pelvic nerve completely abolished both the inhibitory effect upon the reflex EMG activity of the ilio- and pubococcygeus muscles and the facilitation of the iliacus and psoas major muscles provoked by cervical stimulation. Combined pudendal and genitofemoral nerve transection abolished the ilio- and pubococcygeus muscles activation evoked by clitoral and perigenital skin mechanostimulation, but did not affected the iliacus and psoas major muscles activation produced by cervical stimulation with the glass probe (Fig. 7).
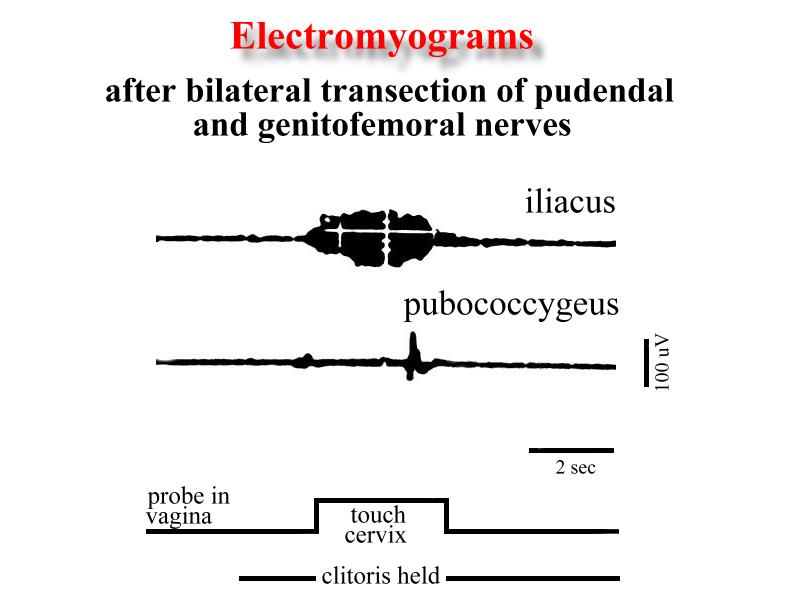 Click to enlarge
Fig. 7
Click to enlarge
Fig. 7
Bilateral transection of either the pudendal or the genitofemoral nerves did not block the ilio- and pubococcygeus muscles activation that was produced by clitoral or perigenital skin or distal vagina stimulation.
Experiment B. It was found that the external vaginal orifice is usually closed. However, ones on a while dilates ("spontaneously"). Tapping of the perigenital area provokes VODs, but not all taps evoke this response. Thus, the proportion of both spontaneous or provoked VODs is depending of the estrous stage (Fig. 8) and of the endocrine condition (Fig. 9) of the female. Fig. 8 shows that taps are proportionally effective according to the different stages of the estrous cycle, being estrous the more significant. Proestrous and estrous were the stages in which more spontaneous VODs were recorded.
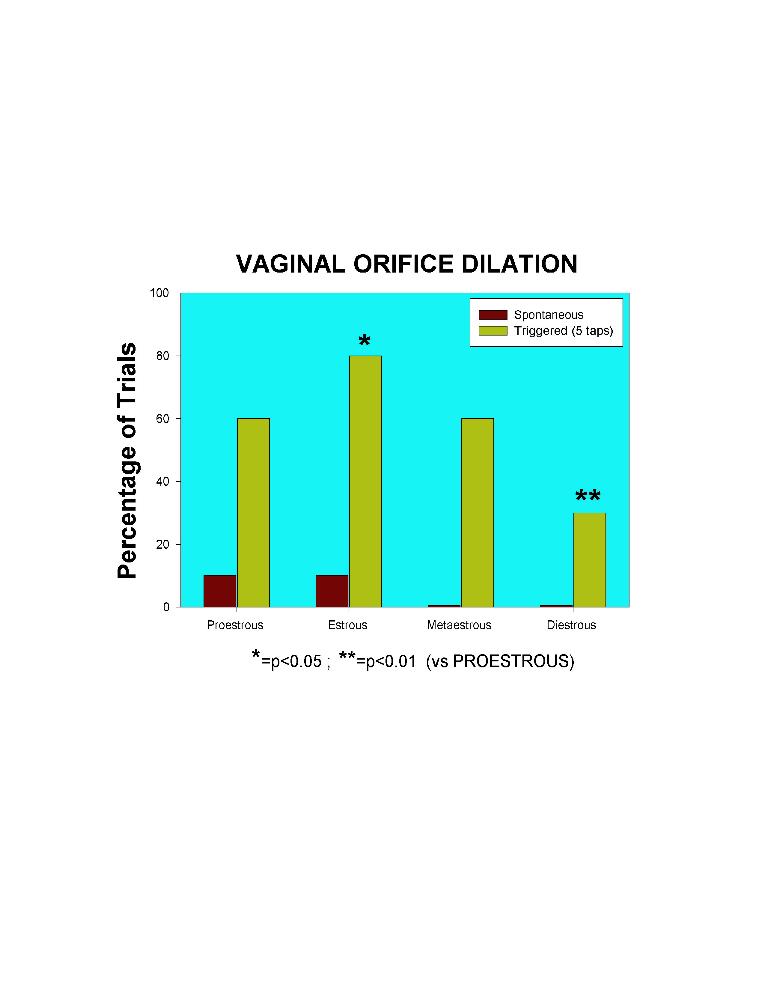 Click to enlarge
Fig. 8
Click to enlarge
Fig. 8
Fig. 9 shows that Ovx abolishes the spontaneous VODs and reduced the effectiveness of tapping. Estradiol benzoate alone or in combination with progesterone restores both taping efectiveness and presence of spontaneous VODs.
 Click to enlarge
Fig. 9
Click to enlarge
Fig. 9
To probe that the VODs observed after tapping was a reflexive reaction, bilateral transection of pelvic, genitofemoral and pudendal nerves were done in Ovx rats. Fig. 10 shows that two weeks after transection of the pelvic nerve there is no effect on VODs provoked in trials of 20 taps. Transection of the genitofemoral nerve reduces on 15% the effectiveness of tapping. The pudendal nerve transection reduces on 55% the effect of tapping.
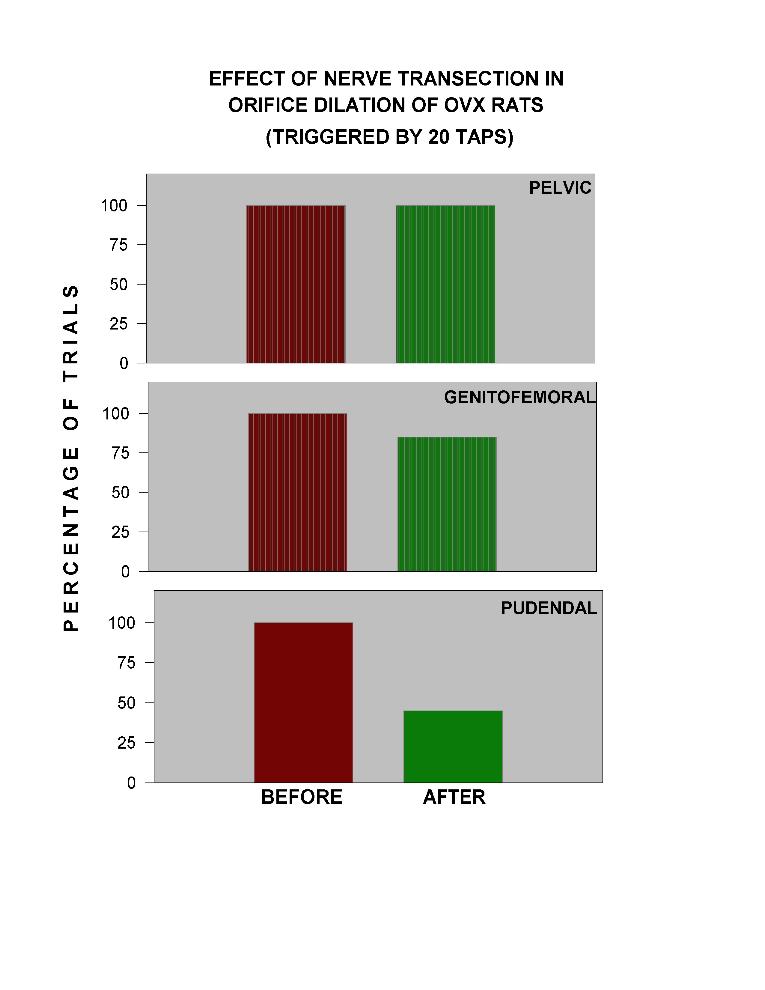 Click to enlarge
Fig. 10
Click to enlarge
Fig. 10
Discussion and Conclusion
Cats The female cat assumes a crouching posture when mounted by the male. To maintain this position and support the male, a strong tonic contraction of the gastrocnemius muscle (GM) among other muscles is needed (Pacheco et al., 1976). In fact, this hypothesis is supported by our findings which showed that the neural circuit involving GM contraction is facilitated by sensory stimulation of the genital skin area (tapping) or by the neckgrip. Furthermore, the lordotic reflex response displayed by an estrous female cat during mating is principally triggered by the mechanical stimulation that the male carries out on specific skin areas, perineum, vagina and cervix. As we show in the present report vaginal stimulation also facilitates the monosynaptic response of the GM. On the other hand, in intact and Ovx animals cervical stimulation produces an inhibitory effect upon the monosynaptic response of the GM. However, in the Ovx animals daily repetitive injections of estrogen facilitates instead the effect of cervical stimulation. This response can be similar in estrous female cats. The tendency for some female cats to maintain lordosis during and after the withdrawal of the glass rod used for artificial stimulation of the urogenital sinus and vagina (Diakow, 1971) correlates with our present results. In summary, the finding that the changes elicited by genital stimulation were dependent of the endocrine condition (intact, Ovx, or Ovx + estrogen), even in acute spinal preparations, agrees with recent observations of direct effects of hormones upon some motoneurons (Kurz et al., 1986). It also indicates that the well known alteration in sexual responsiveness following estrogen administration (Komisaruk, 1978) are due to both action of the hormone at higher nervous levels and low spinal levels. Additionally we want to point out that since the GM is an extensor muscle, our results also show that responses of the skin of the genital area do not accord with what Sherrington (1910) stated as a general rule, that the stimulation of a cutaneous nerve excites the motoneurons of flexor muscles and inhibits the motoneurons of extensor muscles.
Rats Experiment A. Our findings demonstrate differences between the EMG of muscles of the pelvic floor (inhibited) and the hindlimb (activated) in response to the same cervical stimulus. Thus, cervical stimulation elicits reflexive contractions of postural muscles (iliacus and psoas major) but abolishes reflexive contractions of pelvic muscles produced by perineal stimulation. These phenomenae reflects excitatory as well as inhibitory actions upon the corresponding motoneurons. Repeated cervical stimulation elicited sensitization of the reflexive afterdischarge of the hindleg postural muscles recorded. This effect is in accord with the study of Berkeley et al. (1990) upon pelvic nerve afferents. Furthermore, seems that this sensitization is involved in the lordosis posture of female copulation. Therefore, it appears likely that the prolongation of the lordotic posture response (Pfaff et al., 1977; Rodriguez-Sierra et al., 1975) observed in female rats following intromissions and ejaculations (Diakow, 1970, 1975; Komisaruk and Diakow, 1973) or seminal plug deposition (Matthews and Adler, 1977, 1978; Wallach and Hart, 1983) is mediated in part through the above described processes. Inhibition of the pelvic musculature (ilio- and pubococcygeus) also could facilitate withdrawal of the penis from the vagina after ejaculation therby helping to avoid dislodgement of the copulatory plug (Blandau, 1945; Matthews and Adler, 1978; Wallach and Hart, 1983).
Experiment B. Our results show that the branch of the pelvic nerve which innervates the perineal skin of the female rat (Pacheco et al., 1989) does not participates in the VOD reflex. Pudendal nerve which has a rich sensory innervation of the clitoral sheat is the one that plays a major role on the reflex mechanism of the VOD. During the copulatory thrusting the male activates receptors from both pudendal and genitofemoral nerves that reflexively produces VOD facilitating the intromission of the penis. This reflexive reaction seems to be modulated by estradiol and progesterone. Thus, during copulation VOD seems to be triggered through physiological mechanisms of the spinal cord that in harmony with lordosis and contraction of perineal musculature of the pelvic floor facilitates the intromission of the penis. The harmony of these processes is developed with more intensity when the gonadal hormones are present.
References
- Bard, P. (1935) The effects of denervation of the genitalia on the oestral behavior of cats. Am. J. Physiol. 113:5.
- Diakow, C. (1971) Effects of genital desensitization on mating behavior and ovulation in the female cat. Physiol. Behav. 7:47-54.
- Komisaruk, B.R. (1978) The nature of the neural substrate of female sexual behaviour in mammals and its hormonal sensitivity: Review and speculations. In: Hutchinson, J.B. (Ed.). Biological determinants of sexual behavior. John Wiley, New York. pp 349-393.
- Pacheco, P., Beyer, C., Mexicano, G., and Larsson, K. (1976) Effects of genital stimulation upon spinal reflex activity of female cats under various hormonal conditions. Physiol. Behav. 17:699-703.
- Kurz, E.M., Sengelaub, D.R. and Arnold, A.P. (1986) Androgens regulate the dendritic lenght of mammalian motoneurons in adulthood. Science. 232:395-398.
- Sherrington, C.S. (1910) Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. J. Physiol. 40:28-121.
- Pacheco, P., Martínez-Gómez, M., Whipple, B., Beyer, C., and Komisaruk, B.R. (1989) Somato-motor components of the pelvic and pudendal nerves of the female rat. Brain Res. 490:85-94.
- Berkeley, K.J., Hotta, H., Robbins, A., and Sato, Y. (1990) Functional properties of afferent fibers supplying reproductive and other pelvic organs in pelvic nerve of female rats. J. Neurophysiol. 63:256-272.
- Rodriguez-Sierra, J.F., Crowley, W.R., and Komisaruk, B.R. (1975) Vaginal stimulation in rats induces prolonged lordosis responsiveness and sexual receptivity. J. Comp. Physiol. Psychol. 89:79-85.
- Diakow, C. (1970) Effects of genital desensitization on the mating pattern of female rats as determined by motion picture analysis. Am. Zool. 10:486.
- Diakow, C. (1975) Motion picture analysis of rat mating behavior. J. Comp. Physiol. Psychol. 88:704-712.
- Komisaruk B.R. and Diakow, C. (1973) Lordosis reflex intensity in rats in relation to the estrous cycle, ovariectomy, estrogen administration and mating behavior. Endocrinolog. 93:548-557.
- Matthews, M.K. and Adler, N.T. (1977) Facilitative and inhibitory influences of reproductive behavior on sperm transport in rats. J. Comp. Physiol. Psychol. 91:727-741.
- Matthews, M.K. and Adler, N.T. (1978) Systematic interrelationship of mating, vaginal plug position, and sperm transport in the rat. Physiol. Behav. 20:303-309.
- Wallach, S.J.R. and Hart, B.L. (1983) The role of the striated penile muscles of the male rat in seminal plug dislodgement and deposition. Physiol. Behav. 31:815-821.
- Blandau, R.J. (1945) On the factors involved in sperm transport through the cervix uteri of the albino rat. Am. J. Anat. 77:253-272.
- Pfaff, D.W., Montgomery, M., and Lewis, C. (1977) Somatosensory determination of lordosis in female rats. Behavioral definition of the estrogen effect. J. Comp. Physiol. Psychol. 91:134-145.
| Discussion Board | Previous Page | Your Symposium |