Invited Symposium: Genital Sensation: CNS Targets and Functions in Females
| INABIS '98 Home Page | Your Session | Symposia & Poster Sessions | Plenary Sessions | Exhibitors' Foyer | Personal Itinerary | New Search |
Text
INTRODUCTION: Effects of vaginocervical stimulation (VS). Neurally-mediated effects of reproductive tract stimulation, at least in the female rat, can be grouped into three main classes: 1) behavioral , including induction of sexual receptivity, intensification of the lordosis response, immobilization, and analgesia, 2) neuroendocrine , including release from the pituitary of oxytocin, LH and LTH, and 3) autonomic , including net sympathetic activation indicated by increased heart rate, blood pressure and pupil dilatation (for review, see 29 and 38).
A VS-produced neuroendocrine reflex: oxytocin release into plasma In a classical neuroendocrine reflex, the Ferguson reflex (21), distension of the birth canal as the fetus emerges releases oxytocin stored in the posterior pituitary into the systemic circulation, and the oxytocin stimulates uterine contractions, accelerating the delivery of the young (22, review: 35). Plasma oxytocin is also elevated in response to vaginal stimulation during mating (56). Oxytocin facilitates sexual receptivity (3), probably by stimulating the reproductive tract, for deafferentation of the reproductive tract blocks the receptivity-facilitating effect of oxytocin (42) (Fig. 1) .
 Click to enlarge
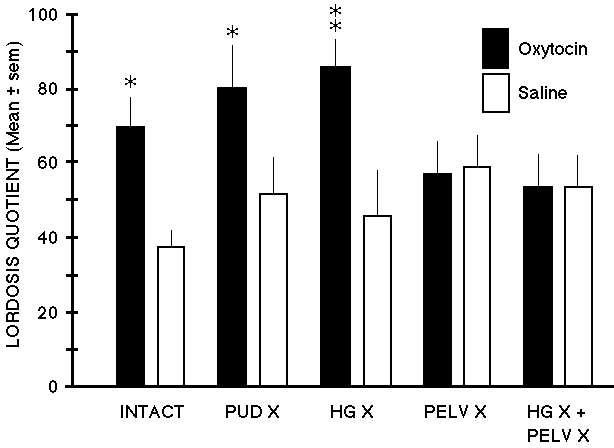
Fig. 1: Oxytocin injected systemically stimulates sexual receptivity, but not if the reproductive tract is denervated. In estrogen-progesterone primed ovariectomized rats, systemic administration of oxytocin (2.1ug) significantly increased the lordosis quotient in nerve-intact control (INTACT), pudendal-neurectomized (PUD X), and hypogastric-neurectomized (HG X) rats. However, pelvic-neurectomized (PELV X) and combined-neurectomized (HG X + PELV X) rats failed to respond to oxytocin.
The pudendal nerves provide sensory innervation of predominantly the external peri- genital skin and clitoral region; the hypogastric nerves provide sensory innervation of predominantly the uterus and cervix. The pelvic nerves provide sensory innervation predominantly of the vagina and cervix. The findings suggest that oxytocin acted peripherally rather than on the central nervous system to stimulate contractions of the reproductive tract and thereby generate sensory input that facilitated receptivity. Data adapted from Moody, et al (1994).
Click to enlarge
Fig. 1: Oxytocin injected systemically stimulates sexual receptivity, but not if the reproductive tract is denervated. In estrogen-progesterone primed ovariectomized rats, systemic administration of oxytocin (2.1ug) significantly increased the lordosis quotient in nerve-intact control (INTACT), pudendal-neurectomized (PUD X), and hypogastric-neurectomized (HG X) rats. However, pelvic-neurectomized (PELV X) and combined-neurectomized (HG X + PELV X) rats failed to respond to oxytocin.
The pudendal nerves provide sensory innervation of predominantly the external peri- genital skin and clitoral region; the hypogastric nerves provide sensory innervation of predominantly the uterus and cervix. The pelvic nerves provide sensory innervation predominantly of the vagina and cervix. The findings suggest that oxytocin acted peripherally rather than on the central nervous system to stimulate contractions of the reproductive tract and thereby generate sensory input that facilitated receptivity. Data adapted from Moody, et al (1994).
The source of the oxytocin in the systemic circulation is the neurosecretory neurons primarily of the paraventricular nucleus of the hypothalamus (15,27,28). Recently, these neurons have also been shown to project into the spinal cord (19,24,26,40,59). Some of these neurons have been shown to bifurcate, distributing terminals to both the posterior pituitary and the spinal cord (53).
VS also releases oxytocin into the spinal cord We asked the question: in the rat, does reproductive tract stimulation release oxytocin into the spinal cord, and if so, does that oxytocin produce any of the neurally-mediated effects of reproductive tract stimulation? In earlier studies, reproductive tract stimulation in the ewe had been shown to release oxytocin into plasma and spinal cord (28), and oxytocin administered to the spinal cord in rats had been shown to increase heart rate (61), which is one of the effects of vaginocervical stimulation in the rat (11)
To address the question of whether oxytocin is released into the spinal cord in response to reproductive tract stimulation in the rat, we assayed superfusates of the spinal cord for oxytocin using RIA before, during and after applying vaginocervical mechanostimulation (VS) (50). We observed a significant increase within 5 minutes of the onset of VS. Concurrently, plasma oxytocin was elevated. To rule out the possibility that the oxytocin in the spinal cord superfusate had diffused from the plasma, we repeated the experiment using hypophysectomized rats, and found a comparable increase in oxytocin in the spinal cord superfusate, but not in the plasma (50). Thus, VS evidently releases oxytocin directly into the spinal cord in the rat. This response is potentiated by estrogen priming (23).
What is the function of VS-produced oxytocin release into spinal cord? To address this question, we microinjected oxytocin directly to the spinal cord via catheter chronically-implanted into the subarachnoid space (i.e. intrathecally: �IT�) at the thoraco-lumbar level of the spinal cord, and ascertained its effects through the use of a battery of behavioral and autonomic tests. The oxytocin exerted no measurable effects on lordosis, locomotion, pain threshold (tail flick test), or heart rate. However, blood pressure and pupil diameter were significantly elevated in response to 10ug oxytocin IT with a latency of less than one minute (50)
VS-released oxytocin into spinal cord produces pupil dilatation This finding raised the question of whether the oxytocin released into the spinal cord in response to VS is the mediator of the pupil dilatation and/or increase in blood pressure response to VS. To test this, we administered a selective oxytocin receptor antagonist (OTA) (25ug in 5ul saline, IT) and in its presence, ascertained the effect of VS on pupil diameter and blood pressure. This OTA significantly and markedly reduced the pupil dilatation response to VS. The blood pressure response to VS was not affected by the OTA. The lack of effect on blood pressure indicates that for this response either there is a different subtype of oxytocin receptor for blood pressure, or that VS releases an additional, non-oxytocic substance that increases blood pressure. The finding that the OTA significantly attenuated the pupil dilatation response to VS, is the most conclusive evidence that the oxytocin that is released into the spinal cord in response to vaginocervical stimulation produces pupil dilatation (50).
Neural pathway for VS-produced release of oxytocin into spinal cord By which neural pathway does the oxytocin dilate the pupil? Since the latency of effect of the oxytocin and the oxytocin antagonist are each less than one minute, it is most likely that the site of action of the oxytocin is in the spinal cord per se; it takes substantially longer for an IT injection to diffuse to the brain (60). We can not rule out the possibility that the oxytocin stimulates an ascending pathway to the oculomotor nucleus in the midbrain, inhibition of which could produce pupil dilatation. However, it is more likely that the oxytocin stimulates the preganglionic cell bodies of the sympathetic division of the autonomic system that are located at spinal cord thoracic levels 1-3 (2) . The postganglionic neurons to which these project are located in the superior cervical ganglion. The postganglionics project directly to the ciliary muscle of the iris and when stimulated, dilate the pupil (2). We hypothesized that the oxytocin released by VS in the spinal cord stimulates these sympathetic preganglionic cell bodies and thereby dilates the pupil.
Visualization of VS-produced oxytocin release into spinal cord To test this hypothesis, we ascertained whether oxytocin can be visualized at the pregangionic neuronal soma in the spinal cord, and whether VS produces an observable change in that oxytocin. The pupillary response is measured using a surgical microscope with a reticle, and observing the pupil of the eye of a rat that is gently restrained (wrapped in a towel), using low intensity illumination. When VS is applied, there is an immediate dilatation of the pupil (50,54). The VS group contained significantly more oxytocin-stained cells than the control group.
Oxytocin: a CNS autonomic sympathetic neurotransmitter? The above findings suggest that oxytocin is an endogenous CNS neurotransmitter for the sympathetic division of the autonomic nervous system. We also conclude that the pupil dilatation response to VS is mediated at least in large part by this oxytocin. These conclusions are based on the following; in summary : a) VS produces pupil dilatation, b) VS releases oxytocin into the spinal cord, c) oxytocin administration to the spinal cord produces pupil dilatation, d) an oxytocin receptor blocking agent markedly attenuates the pupil dilatation that is produced by VS, e)VS releases oxytocin-like immunoreactivity in the vicinity of autonomic, sympathetic-division preganglionic neurons. In addition, paraventricular oxytocinergic neurons have been shown to project to sympathetic preganglionic neurons in the spinal cord (49), iontophoretic application of oxytocin to sympathetic preganglionic neurons in the spinal cord alters their firing rate (24,41, for review: 47), and oxytocin administration to the spinal cord increases heart rate (4,61).
Afferent pathway(s) by which VS accesses the oxytocinergic neurons . On the basis of studies in which axonal recording was used to map sensory fields of the reproductive system, the pudendal nerves provide sensory innervation predominantly of the skin surrounding the vaginal orifice and clitoris (30), the hypogastric nerves provide sensory innervation predominantly of the cervix and uterus (6, 7, 8,46), and the pelvic nerves provide sensory innervation predominantly of the vagina and cervix (6, 7, 8,30,46). Surgical transection of the pelvic and hypogastric nerves in rats significantly attenuates VS-produced analgesia (18), the analgesia of mating (25), the female's pacing of intromissions (20), ovulation produced by uterine stimulation (17), and mating-induced hormonal responses necessary for pregnancy (1,55). The pelvic nerves project to the spinal cord at levels L6-S1, whereas the hypogastric nerves project to the spinal cord at more rostral levels, i.e. T13-L4 16,52). While both pairs of nerves convey cervical sensory activity to the spinal cord, only the pelvic nerve projects to the sacral level of the spinal cord.
A VS-activated spino-hypothalamic pathway There is evidence, based on use of the retrograde tracer, Fluorogold, of the existence of a spino-hypothalamic pathway from sacral spinal cord to various sites in the hypothalamus, including the paraventricular nucleus (10). Based on use of c-fos immunostaining as a marker of neuronal cell bodies that respond to sensory stimulation,we found sites in the sacral spinal cord that respond to VS (13). Some of the sites in the sacral spinal cord in these two independent studies (10,13) appear closely congruent. On the basis of this apparent congruency, we hypothesized that there exist neurons in the spino-hypothalamic tract projection to the paraventricular nucleus that respond to VS.
To test this hypothesis, we used a double-labeling method in which we injected Fluorogold into the paraventricular nucleus and a week later, applied VS, waited 2h and then euthanized the rats. We observed at least 2 double-labelled cells in the sacral spinal cord. This indicates that there exist neurons in the sacral spinal cord that respond to VS and project directly to the paraventricular nucleus of the hypothalamus (12). It is probable that these spino-hypothalamic neurons are second-order neurons that receive input directly from the pelvic nerve.
A 3-neuron pathway for the Ferguson Reflex? Based on the above findings, we propose that the simplest (i.e. fewest neurons) pathway for the Ferguson reflex is: 1 �: pelvic nerve afferents, 2 �: sacral spino-hypothalamic neuron, 3 �:oxytocin-synthesizing neurosecretory neuron of the paraventricular nucleus that projects to the neural lobe of the pituitary gland. Thus, the mechanostimulation of the reproductive tract during parturition could release oxytocin into the systemic circulation via a pathway that contains a series of as few as three neurons.
A 5-neuron pathway for VS-produced pupil dilatation? Extrapolating this pathway to the pupil dilatation response to VS, third order (3 �) oxytocin-synthesizing neurons in the paraventricular nucleus could project down the spinal cord to the the intermediolateral column at T1-3 where they synapse onto the 4o sympathetic preganglionic neurons containing oxytocin receptors, that project into the sympathetic chain gangia and up to the superior cervical ganglion, where they synapse onto the 5o sympathetic postganglionic neurons that project to the radial muscle of the pupil, activation of which produces pupil dilatation. Thus, the neural pathway for the oxytocin-mediated, sympathetic division-mediated, pupil-dilatation response to VS contains a sequence of as few as five neurons
Evidence of a genital sensory pathway that bypasses the spinal cord Although the sensory fields of the pelvic and hypogastric nerves overlap in the cervix, they differentially innervate the vagina and uterus (for review: 38), differ in the regions of the body to which they provide optimal analgesia (36), and project to different levels of the spinal cord (52). Since in humans the hypogastric nerve projects to the spinal cord levels T10-T12 (9,43), we hypothesized that significant genital sensory activity could be perceived by women with complete spinal cord injury at the relatively high level of T11, since sensory input would still be able to enter the spinal cord at T10, even though input via the pelvic nerve projection to the spinal cord would be interrupted (37,34). There are anecdotal reports of sexual response in women with complete spinal cord injury at these relatively high levels (5,14). Furthermore, we hypothesized that women with complete spinal cord injury above T10 should not be able to perceive sensory input from the reproductive tract since all the known genitospinal nerves (i.e. pudendal, pelvic and hypogastric) enter the spinal cord at or below this level. To test this hypothesis, we performed a study of perceptual responses to vaginal and cervical self-stimulation in women diagnosed with complete spinal cord injury at or above T10, which should block all genital sensory perceptual responses compared to women with complete spinal cord injury below T10, which should permit at least some genital sensory perceptual responses. A control group comprised women without spinal cord injury (37,34).
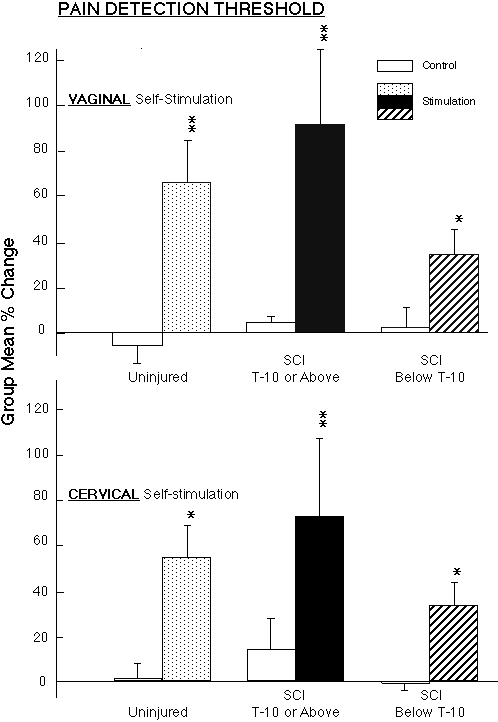
Genital sensory response in women with "complete" spinal cord injury In support of the hypothesis, the women in the group with the lower level of spinal cord injury did show significant analgesia, measured at the fingers, in response to vaginal or cervical self-stimulation (Fig. 2). To our surprise, the women with the upper level of spinal cord injury, who we had expected would not respond, showed the greatest magnitude analgesic response to vaginal and cervical self-stimulation of the three groups. In addition, all the women in both spinal cord injury groups reported that they experience menstrual discomfort or cramps, some women in each of the spinal cord injury groups reported vaginal and/or cervical sensory awareness, and at least one woman in each of the spinal cord injury groups experienced orgasms from the vaginal and/or cervical self-stimulation during the laboratory study (34,57). (The findings of orgasmic response in women with complete spinal cord injury at these levels are supported by another recent study: 51).
 Click to enlarge
Fig. 2: Vaginal and Cervical self-stimulation produced significant increases in mean ( � sem) pain detection threshold (measured by applying pressure to the fingers) in non-injured women (n=5), in women with complete spinal cord injury below T-10 (n=10) (above the level of entry into the spinal cord of the pudendal and pelvic nerves, but not all components of the hypogastric nerve), and in women with complete spinal cord injury at T-10 or above (n=6) (above the level of entry into the spinal cord of the pudendal, pelvic and hypogastric nerves). Within-group paired comparisons: ** p<0.01, *p<0.05: during-stimulation levels compared to baseline control levels.
Click to enlarge
Fig. 2: Vaginal and Cervical self-stimulation produced significant increases in mean ( � sem) pain detection threshold (measured by applying pressure to the fingers) in non-injured women (n=5), in women with complete spinal cord injury below T-10 (n=10) (above the level of entry into the spinal cord of the pudendal and pelvic nerves, but not all components of the hypogastric nerve), and in women with complete spinal cord injury at T-10 or above (n=6) (above the level of entry into the spinal cord of the pudendal, pelvic and hypogastric nerves). Within-group paired comparisons: ** p<0.01, *p<0.05: during-stimulation levels compared to baseline control levels.
Evidence that the Vagus nerve is a genital sensory pathway to the brain Which afferent neural pathway could account for the findings of perceptual responses to genital self-stimulation in women with complete spinal cord injury above the level of entry into the spinal cord of all the known genitospinal nerves? In 1990, it was reported in rats that the vagus nerve (Cranial Nerve #10) innervates the reproductive tract, on the basis that injection of the marker, horseradish peroxidase, into the walls of the cervix and uterus labeled neurons in the nodose ganglion, which is the dorsal root ganglion of the vagus nerve (45). This suggested a possible pathway to account for the responsiveness in the women with spinal cord injury. We tested the functionality of this pathway in rats by surgically transecting the spinal cord at T7, ascertaining whether any brain-mediated responses to VS persisted, and then transected the vagus nerves bilaterally and retested the rats for responses to VS (31). In a second study, we transected the pelvic, hypogastric and pudendal nerves bilaterally, tested for residual brain-mediated responses to VS, then transected the vagus nerves bilaterally and retested the rats for responses to VS (16). In both studies, after genitospinal nerve or spinal cord transection, VS still produced significant pupil dilatation, albeit lower in magnitude. Subsequent vagotomy abolished the response. Residual changes in pain threshold to VS after genitospinal nerve transection or spinal cord transection were also abolished following subsequent vagotomy (16,31).
Vagal nerve afferent stimulation produces pupil dilatation Since these studies showed that the pupil dilatation response to VS is evidently mediated in part by the vagus nerves, we hypothesized that electrical stimulation of the nerve would mimic this effect. To test this hypothesis, we transected a vagus nerve, applied electrical stimulation to the central stump of the nerve, and measured pupil diameter. We observed immediate and significant pupil dilatation, the magnitude of which was directly proportional to the applied voltage, thus supporting the hypothesis (33). To further characterize the pathway, we ascertained whether the sympathetic innervation of the eye mediates this pupil dilatation response to vagal stimulation. We considered the possibility that the NTS projects down to the spinal cord to the sympathetic pre- and post-ganglionics to the pupil. To test this, we surgically ablated bilaterally the superior cervical ganglion plus several of the first sympathetic chain ganglia, then repeated the electrical stimulation of a central stump of the transected vagus nerve. The pupil dilatation response to the vagal stimulation persisted unabated (33). This ruled out a pathway via the sympathetic innervation of the pupil.
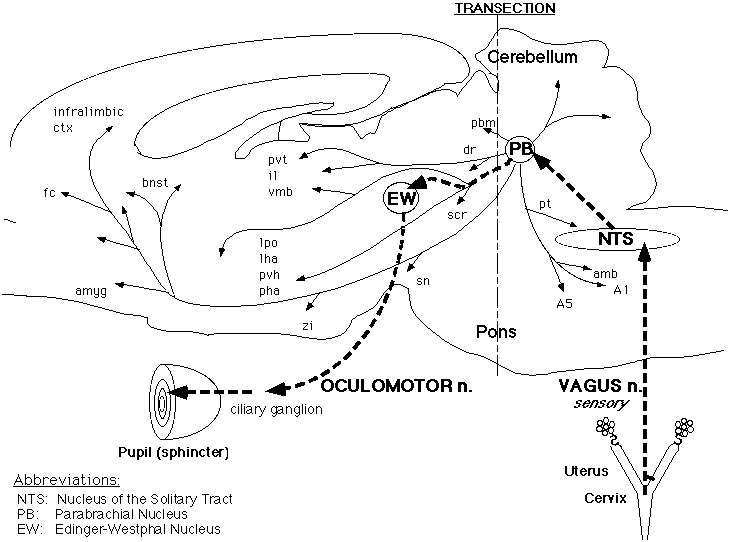
A vagal afferent parasympathetic-inhibitory pathway? Our alternate hypothesis is that the NTS projects rostrally to the oculomotor nucleus of the midbrain, which provides the parasympathetic innervation of the pupil. A possible pathway from the NTS is to the parabrachial nucleus, which projects to the oculomotor nucleus. In order to test this hypothesis, we first obtained pupil dilatation to electrical stimulation of the central stump of the transected vagus nerve, then performed a mid-pontine transection using a micro-knife. In the results of a pilot study, the midpontine transection abolished the response to vagal stimulation. Moreover, the pupil constriction response to shining a light into the eye persisted. This indicates that the final common pathway to the pupil remained intact after the midpontine transection, although the response to vagal stimulation was abolished (Fig. 3) (33). On the basis of the results of this study to date, vagal afferent activity evidently produces pupil dilatation not by stimulation of the sympathetic innervation, but instead by inhibition of the parasympathetic innervation of the pupil.
 Click to enlarge
Fig. 3: Schematic representation of the putative pathway by which the Vagus nerve mediates pupil dilatation in response to vaginocervical stimulation in the rat. Electrical stimulation of the central stump of the Vagus nerve produced pupil dilatation, which was not affected by bilateral ablation of the Superior Cervical Ganglia (SCG), thus ruling out a pathway via descending connections to the SCG from the Nucleus of the Solitary Tract (NTS) (to which the the Vagal primary afferents project). Subsequent total transection of the brainstem at the midpontine level abolished the pupil dilatation response to Vagal stimulation, but did not block the pupillary light reflex, indicating that the final common pathway to the pupil was still functional. The major putative relays in this genital sensory pathway are: NTS to Parabrachial Nucleus (PB) to Edinger-Westphal nucleus (EW) via the oculomotor nerve to the ciliary ganglion. Since activation of this parasympathetic nerve produces pupillary constriction, we propose that stimulation of the Vagal afferents must inhibit the oculomotor output, at one or more of these relays.
Click to enlarge
Fig. 3: Schematic representation of the putative pathway by which the Vagus nerve mediates pupil dilatation in response to vaginocervical stimulation in the rat. Electrical stimulation of the central stump of the Vagus nerve produced pupil dilatation, which was not affected by bilateral ablation of the Superior Cervical Ganglia (SCG), thus ruling out a pathway via descending connections to the SCG from the Nucleus of the Solitary Tract (NTS) (to which the the Vagal primary afferents project). Subsequent total transection of the brainstem at the midpontine level abolished the pupil dilatation response to Vagal stimulation, but did not block the pupillary light reflex, indicating that the final common pathway to the pupil was still functional. The major putative relays in this genital sensory pathway are: NTS to Parabrachial Nucleus (PB) to Edinger-Westphal nucleus (EW) via the oculomotor nerve to the ciliary ganglion. Since activation of this parasympathetic nerve produces pupillary constriction, we propose that stimulation of the Vagal afferents must inhibit the oculomotor output, at one or more of these relays.
The genital-Vagal hypothesis in women: Preliminary P.E.T. scan analysis These studies support the hypothesis that the vagus nerve can convey afferent activity from the reproductive tract directly to the medulla oblongata of the brain, bypassing the spinal cord. To test whether this is a functional pathway that could account for our findings in women with complete spinal cord injury, we have begun a study using Positron Emission Tomography (PET) to ascertain whether cervical self-stimulation in women with complete spinal cord injury activates the region of the Nucleus of the Solitary Tract (NTS), which receives the primary afferent terminals of the vagus nerve. In a pilot study with two women with complete spinal cord injury above T-10, we have observed evidence of activation of the region of the NTS in response to cervical self-stimulation (39).
Is Vagal afferent stimulation adequate to produce orgasm? The evident genital sensory role of the vagus nerve is significant from a clinical and a theoretical perspective. Clinically, the vagus nerve may provide a previously unrecognized spinal cord bypass channel from the genital system, whose activity could be augmented by pharmacological and/or biofeedback methods, a procedure that might be able to potentiate sexual response. From a theoretical perspective, since the vagus nerve evidently conveys genital afferent activity and thereby can mediate sexual responses, it provides a localized, specific pathway by which to analyze the neural and neurochemical mechanism underlying sexual response and orgasm, eliminating the experimental complexity of assessing the role of the spinal cord input to the brain.
Finally, the question remains unanswered as to the functional role, if any, of the pupil dilatation response to VS. This is a robust and reliable response that has redundant neural representation, both via the spinal cord and via the vagus nerves. Pupil dilatation occurs normally in response to intromittive stimulation in rats (54) and in humans (58). Whether it plays a significant role in social signalling and/or adaptation (e.g. visual accommodation) to the conditions of close social proximity during sexual behavior, is an open and intriguing question.
References
1.- Adler, N.T., Resko, J.A., Goy, R.W. The effect of copulatory behavior on hormonal change in the female rat prior to implantation. Physiology and Behavior. 5:1003 1007. 1970.
2.-Appenzeller, O. The Autonomic Nervous System. An introduction to basic and clinical concepts 4th ed. Elsevier. New York. 1990.
3.- Arletti, R., Bertolini, A. Oxytocin stimulates lordosis behavior in female rats. Neuropeptides. 6:247-253. 1985.
4.- Backman, S.B., Henry, J.L. Effects of oxytocin and vasopressin on thoracic sympathetic preganglionic neurons in the cat. Brain Res. Bull. 13:679-684. 1984.
5.- Berard, E.J.J. The sexuality of spinal cord injured women: Physiology and pathophysiology: A review. Paraplegia. 27:99-112. 1989.
6.- Berkley, K.J., Hotta, H., Robbins, A., Sato, Y. Functional properties of afferent fibers supplying reproductive and other pelvic organs in pelvic nerve of female rats. Journal of Neurophysiology. 63:256-272. 1990.
7.- Berkley, K.J., Robbins, A., Sato, Y. Afferent fibers supplying the uterus in the rat. J. Neurophysiol. 59:142-163. 1988.
8.- Berkley, K.J., Robbins, A., Sato, Y. Functional differences between afferent fibers in the hypogastric and pelvic nerves innervating female reproductive organs in the rat. J. Neurophysiol. 69:533-544. 1993.
9.-Bonica, J.J. Principles and Practices of Obstetric Analgesia and Anesthesia. F.A. Davis Company. Philadelphia. 1967.
10.- Burstein, R., Cliffer, K.D., Giesler, G.J. Cells of origin of the spinohypothalamic tract in the rat. Journal of Comparative Neurology. 291:329-344. 1987.
11.- Catelli, J.J., Sved, A.F., Komisaruk, B.R. Vaginocervical stimulation elevates blood pressure and induces analgesia by separate mechanisms. Physiology and Behavior. 41:609-612. 1987.
12.-Chinapen, S. Aging-related reduction in responsiveness to vaginocervical stimulation in rats: behavioral and neuroanatomical analysis. Doctoral dissertation. Rutgers - The State University of New Jersey. 1998.
13.- Chinapen, S., Swann, J.M., Steinman, J.L., Komisaruk, B.R. Expression of c fos protein in lumbosacral spinal cord in response to vaginocervical stimulation in rats. Neuroscience Letters. 145:93-96. 1992.
14.- Cole, T. Sexuality and physical disabilities. Arch Sex Behav. 4:389-403. 1975.
15.- Coombes, J., Robinson, I.C.A.F., Antoni, F.A., Russell, J.A. Release of oxytocin into blood and into cerebrospinal fluid induced by naloxone in anaesthetized morphine-dependent rats: the role of the paraventricular nucleus. J. Neuroendocrinol. 3:551-561. 1991.
16.- Cueva-Rolon, R., Sansone, G., Bianca, R., Gomez, L.E., Beyer, C., Whipple, B., Komisaruk, B.R. Vagotomy blocks responses to vaginocervical stimulation in genitospinal-neurectomized rats. Physiol. Behav. 60:19-24. 1996.
17.- Cunningham, S.T., Rosenblatt, J.S., Komisaruk, B.R. Reflexive ovulation in the rat, induced by caesarian-section, is blocked by pelvic and/or hypogastric nerve transection. Neuroendocrinology. 56:393-396. 1992.
18.- Cunningham, S.T., Steinman, J.L., Whipple, B., Mayer, A.D., Komisaruk, B.R. Differential roles of hypogastric and pelvic nerves in the analgesic and motoric effects of vaginocervical stimulation in rats. Brain Research. 559:337-343. 1991.
19.- Dogterom, J., Van Wimersma Greidanus, T.B., Swaab, D.F. Evidence for release of vasopressin and oxytocin into cerebrospinal fluid: measurements in plasma and CSF of intact and hypophysectomized rats. Neuroendocrinology. 24:108-118. 1977.
20.- Erskine, M.S. Pelvic and pudendal nerves influence the display of paced mating behavior in response to estrogen and progesterone in the female rat. Behavioral Neuroscience. 106:690-697. 1992.
21.- Ferguson, J.K.W. A study of the motility of the intact uterus at term. Surg. Gynecol. Obstet. 359-366. 1941.
22.- Flint, A.P.F., Forsling, M.L., Mitchell, M.D., Turnbull, A.C. Temporal relationship between changes in oxytocin and prostaglandin F levels in response to vaginal distension in the pregnant and puerperal ewe. J Reprod. Fertil. 43:551-554. 1975.
23.- Gerdes, C.A., Sansone, G.R., Steinman, J.L., Winslow, J.T., Ottenweller, J.E., Komisaruk, B.R., Insel, T.R. Potentiation of oxytocin release into the spinal cord by estrogen priming in the rat. submitted.:.
24.- Gilbey, M.P., Coote, J.H., Fleetwood-Walker, S., Peterson, D.F. The influence of the paraventriculo-spinal pathway, and oxytocin and vasopressin on sympathetic preganglionic neurons. Brain Res. 251:283-290. 1982.
25.- Gomora, P., Beyer, C., Gonzalez-Mariscal, G., Komisaruk, B.R. Momentary analgesia produced by copulation in female rats. Brain Research. 656:52-58. 1994.
26.- Hawthorn, J., Ang V.T.Y., J.S., J. Effects of lesions in the hypothalamic paraventricular, supraoptic and suprachiasmatic nuclei on vasopressin and oxytocin in rat brain and spinal cord. Brain Res. 346:51-57. 1985.
27.- Higuchi, T., Uchide, K., Honda, K., Negoro, H. Oxytocin release during parturition in the pelvic-neurectomized rat. J. Endocrinol. 109:149-154. 1986.
28.- Kendrick, K.M., Keverne, E.B., Hinton, M.R., Goode, J.A. Cerebrospinal fluid and plasma concentrations of oxytocin and vasopressin during parturition and vaginocervical stimulation in the sheep. Brain. Res. Bull. 26:803-807. 1991.
29.- Komisaruk, B.R. Vaginocervical afference as a trigger for analgesic, behavioral, autonomic and neuroendocrine processes. In:Behavioral Biology: Neuroendocrine Axis. T. Archer and S. Hansen (Eds). Lawrence Erlbaum Associates. Hillsdale, NJ. 1990. pp. 123-137.
30.- Komisaruk, B.R., Adler, N.T., Hutchison, J. Genital sensory field: enlargement by estrogen treatment in female rats. Science. 178:1295-1298. 1972.
31.- Komisaruk, B.R., Bianca, R., Sansone, G., Gomez, L.E., Cueva-Rolon, R., Beyer, C., Whipple, B. Brain-mediated responses to vaginocervical stimulation in spinal cord-transected rats: role of the vagus nerves. Brain Research. 708:128-134. 1996.
32.- Komisaruk, B.R., Caba, M., O'Bannon, Pacheco, P. Vaginocervical stimulation releases oxytocin onto preganglionic sympathetic cell bodies in spinal cord of rat: quantitative immunocytochemical evidence. In preparation.
33.- Komisaruk, B.R., Cueva-Rolon, R., Gomez, L., Ganduglia-Pirovano, M., Sansone, G.R., Bianca, R. Vagal afferent electrical stimulation produces pupil dilatation in the rat. S.N. Abst. 21:1156 (abstract). 1995.
34.- Komisaruk, B.R., Gerdes, C., Whipple, B. "Complete" spinal cord injury does not block perceptual responses to genital self-stimulation in women. Arch. Neurol. 54:1513-1520. 1997.
35.- Komisaruk, B.R., Steinman, J.L. Genital stimulation as a trigger for neuroendocrine and behavioral control of reproduction. In:Reproduction: A Behavioral and Neuroendocrine Perspective. B.R. Komisaruk, H. Siegel, M.-F. Cheng and H.H. Feder (Eds). New York Academy of Sciences. New York. 1986. pp. 64-75.
36.- Komisaruk, B.R., Wallman, J. Antinociceptive effects of vaginal stimulation in rats: Neurophysiological and behavioral studies. Brain Research. 137:85-107. 1977.
37.- Komisaruk, B.R., Whipple, B. Complete spinal cord injury does not block perceptual responses to vaginal or cervical self-stimulation in women. Society for Neuroscience Abstracts. 20:961. 1994.
38.- Komisaruk, B.R., Whipple, B. The suppression of pain by genital stimulation in females. Annual Review of Sex Research. 6:151-186. 1995.
39.- Komisaruk, B.R., Whipple, B., Gerdes, C., Harkness, B., Keyes, J.W.J. Brainstem response to cervical self-stimulation: preliminary PET-scan analysis. S.N. Abst. 23:1001 (abstract). 1997.
40.- Lang, R.E., Heil, J., Banten, D., Hermann, K., Rascher, W., Unger, T. Effects of lesions in the paraventricular nucleus of the hypothalamus on vasopressin and oxytocin contents in brainstem and spinal cord of the rat. Brain Res. 260:326-329. 1983.
41.- Luiten, P.G.M., Ter Horst, G.J., Karst, H., Steffens, A.B. The course of paraventricular hypothalamic efferents to autonomic structures in medulla and spinal cord. Brain Res. 329:374-378. 1985.
42.- Moody, K.M., Steinman, J.L., Komisaruk, B.R., Adler, N.T. Pelvic neurectomy blocks oxytocin-facilitated sexual receptivity in rats. Physiology and Behavior. 56:1057-1060. 1994.
43.-Netter, F.H. The Ciba Collection of Medical Illustrations. Nervous System. Part I. Anatomy and Physiology. Ciba Pharmaceutical Company, Summit, NJ. 1986.
44.- O'Bannon, R.T., III, Komisaruk, B.R. Pupil dilatation response to vaginocervical stimulation in the rat: time course of exhaustion and recovery. submitted. .
45.- Ortega-Villalobos, M., Garcia-Bazan, M., Solano-Flores, L.P., Ninomiya Alarcon, J.G., Guevara-Guzman, R., Wayner, M.J. Vagus nerve afferent and efferent innervation of the rat uterus: An electrophysiological and HRP study. Brain Research Bulletin. 25:365-371. 1990.
46.- Peters, L.C., Kristal, M.B., Komisaruk, B.R. Sensory innervation of the external and internal genitalia of the female rat. Brain Research. 408:199-204. 1987.
47.- Richard, P., Moos, F., Freund-Mercier, M. Central effects of oxytocin. Physiological Rev. 71:331-370. 1991.
48.- Rodriguez-Sierra, J.F., Komisaruk, B.R. Estrogen accelerates the recovery of the lordosis response after its exhaustion induced by cervical probing. Hormones and Behavior. 17: 302-307. 1983.
49.- Rousselot, P., Papadopoulos, G., Merighi, A., Poulain, D.A., Theodosis, D.T. Oxytocinergic innervation of the rat spinal cord. An electron microscopic study. Brain Res. 529:178-184. 1990.
50.-Sansone, G. Hormonal, autonomic and behavioral responses to vaginocervical stimulation: role of the spinal cord and vagus nerve in rats. Doctoral dissertation. Rutgers-The State University of New Jersey. 1996.
51.- Sipski, M., Alexander, C., Rosen, R. Orgasm in women with spinal cord injuries: a laboratory-based assessment. Arch. Phys. Med. Rehabil. 76:1097-1102. 1995.
52.- Steinman, J.L., Carlton, S.M., Willis, D.W. The segmental distribution of afferent fibers from the vaginal cervix and hypogastric nerve in rats. Brain Research. 575:25 31. 1992.
53.- Swanson, L.W., Sawchenko, P.E., Wiegand, S.J., Price, J. Separate neurons in the paraventricular nucleus project to the median eminence and to the medulla or spinal cord. Brain Res. 198:190-195. 1980.
54.- Szechtman, H., Adler, N.T., Komisaruk, B.R. Mating induces pupillary dilatation in female rats: role of pelvic nerve. Physiol. Behav. 35:295-301. 1985.
55.- Traurig, H., Saria, A., Lembeck, F. The effects of neonatal capsaicin treatment on growth and subsequent reproductive function in the rat. Arch. Pharmacol. 327:254-259. 1984.
56.- Vandemark, N.L., Hays, R.L. Uterine motility responses to mating. Am. J. Physiol. 170:518 - 521. 1952.
57.- Whipple, B., Gerdes, C.A., Komisaruk, B.R. Sexual response to self-stimulation in women with complete spinal cord injury. J. Sex Research. 33:231-240,. 1996.
58.- Whipple, B., Ogden, G., Komisaruk, B.R. Physiological correlates of imagery induced orgasm in women. Archives of Sexual Behavior. 21:121-133. 1992.
59.- White, D.J., Krause, J.E., McKelvy, J.F. In vivo biosynthesis and transport of oxytocin, vasopressin and neurophysin from the hypothalamus to the spinal cord. Neuroscience. 17:133-140. 1986.
60.- Yaksh, T.L., Rudy, T.A. Chronic catheterization of the spinal subarachnoid space. Physiol. Behav. 17:1031-1036. 1976.
61.- Yashpal, K., Gauthier, S., Henry, J.L. Oxytocin administered intrathecally preferentially increases heart rate rather than arterial pressure in the rat. J. Auton. Nervous Syst. 20:167-178. 1987.
| Discussion Board | Previous Page | Your Symposium |