| INABIS '98 Home Page | Your Symposium | Related Symposia & Posters | Scientific Program | Exhibitors' Foyer | Personal Itinerary | New Search |
Participation of Placental Opioid-Enhancing Factor in Opioid-Modulated Events at Parturition
Endogenous opioids play a significant role in mammalian parturition as well as in the behaviors associated with delivery itself and toward the emerging young. Distention of the uterus, and distention and mechanical stimulation of the vaginal/cervical area are critical stimuli in the positive feedback loop associated with expulsion of the fetus, and in the onset of maternal behavior upon emergence of the fetus [9, 11, 25]. These stimuli are sufficiently "stressful", "painful", or "aversive" so that they occur during a period of elevated endogenous opioid levels and elevated pain threshold [8, and see 12 for review]. Furthermore, the elevation of endogenous opioids in certain brain areas at delivery, namely the ventral tegmental area, is associated with the rapid onset of appropriate maternal caretaking behavior at that time [24]. One might hypothesize that if endogenous opioids facilitate maternal behavior and delivery, that the level of opioid present would be the same for the optimization of both dimensions of parturition. In fact, increasing pain threshold beyond that observed immediately prior to delivery, in rats, by administration of exogenous opioid (morphine), interferes with the onset of proper maternal care [1, 10, 19, 21]. How then is antinociception increased, at parturition, in a way that does not interfere with the expression of proper maternal behavior?
Placenta and amniotic fluid contain a molecule(s) that potentiates the antinociceptive action of opioids. We refer to this substance as POEF, for Placental Opioid-Enhancing Factor. In an extensive series of experiments, we elucidated many of the features of POEF [see 12 for review]. The following is a summary of the findings of the early phase of POEF research:
- Ingestion of either placenta or amniotic fluid produces opioid-analgesia enhancement (amniotic fluid is available to the mother before expulsion of the fetus, whereas placenta is only available afterward)
- POEF enhances pregnancy-mediated analgesia, as well as that produced by footshock, morphine injection, pseudopregnancy, and vaginal/cervical probing
- POEF does not produce antinociception if administered in the absence of opioid-mediated analgesia, despite the opioid content of afterbirth materials
- Enhanced antinociception is detectable in rats using all assays tested (tail-flick, hot-water tail-immersion, formalin test, hot-plate test)
- POEF does not enhance aspirin-induced [13] or nicotine-induced [18] antinociception in naltrexone-treated rats, and is therefore apparently specific for opioid-mediated antinociception
- POEF must be ingested; it is apparently ineffective if injected
- The optimal amounts for enhancement of 3 mg/kg morphine in rats: 1 placenta (500 mg) or .25 ml amniotic fluid.
- Too high a dose of POEF may produce hyperalgesia
- Enhancement in rats is detectable within 5 min after ingestion and lasts 30 - 40 min
- Male rats show enhancement of morphine antinociception after ingestion of placenta
- Human, bovine, and dolphin placenta contain POEF activity (when tested in rats)
- Bovine amniotic fluid contains POEF activity when tested in cows [16]
- Pregnant-rat liver does not contain POEF activity
- Ingested POEF enhances the central antinociceptive action of morphine [5]
- POEF apparently works by activating gut vagal receptors
- Gastric vagotomy blocks the enhancing action of ingested POEF on morphine antinociception [22]
- Pretreatment with famotidine, an H2 histamine-receptor antagonist, to block digestion, does not block the POEF effect [17]
- During delivery, early amniotic-fluid treatment affects tail-flick latency, contractions, and aggression, depending on time and dose of treatment
- Effects on other opioid-mediated phenomena:
- morphine-induced hyperthermia is unaffected
- contralateral circling after unilateral injection of morphine into the ventral tegmentum is inhibited
- morphine analgesia is produced by a normally subthreshold dose in morphine tolerant rats
- the effect of a low dose of morphine on amelioration of withdrawal symptoms is enhanced
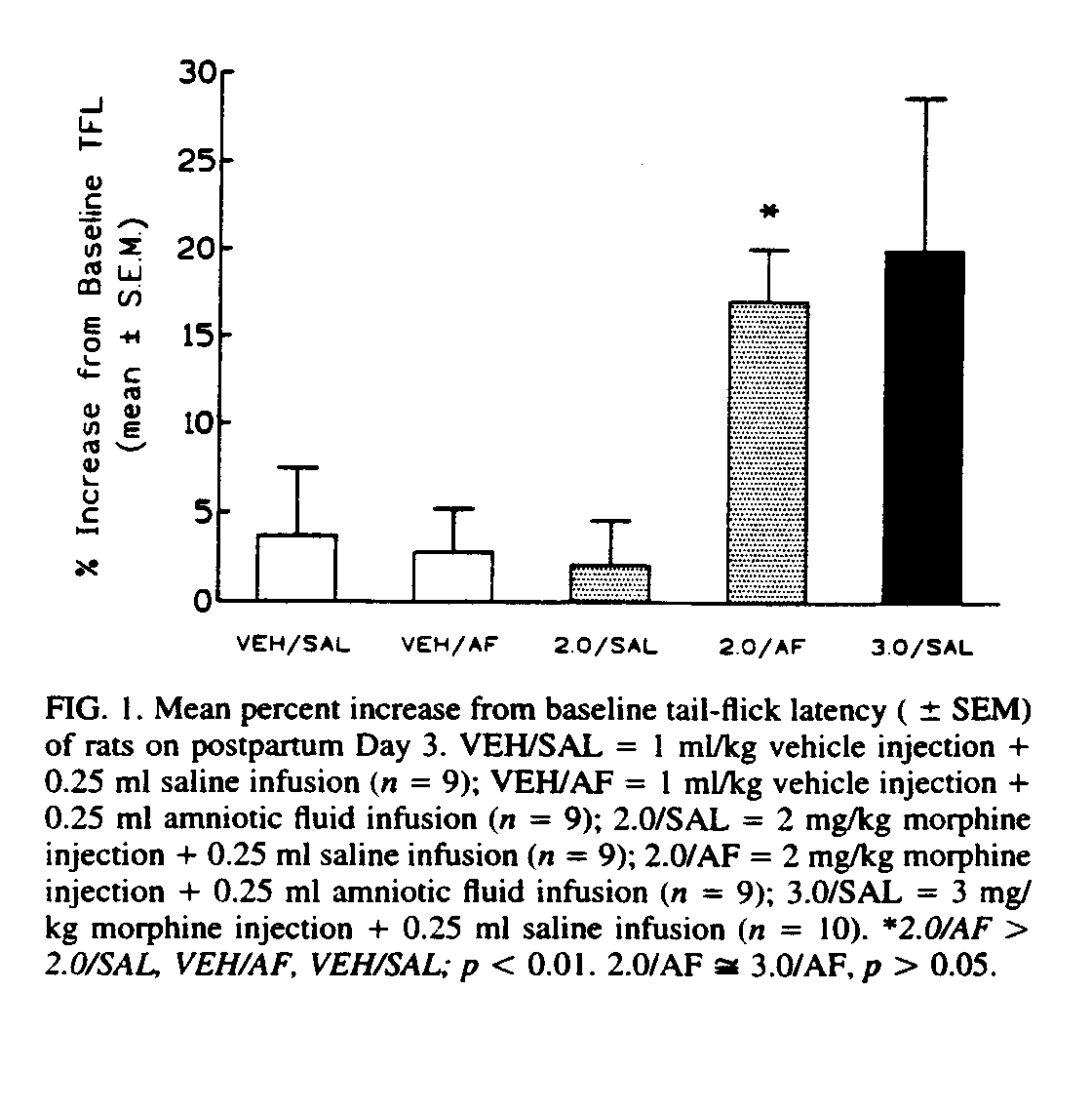
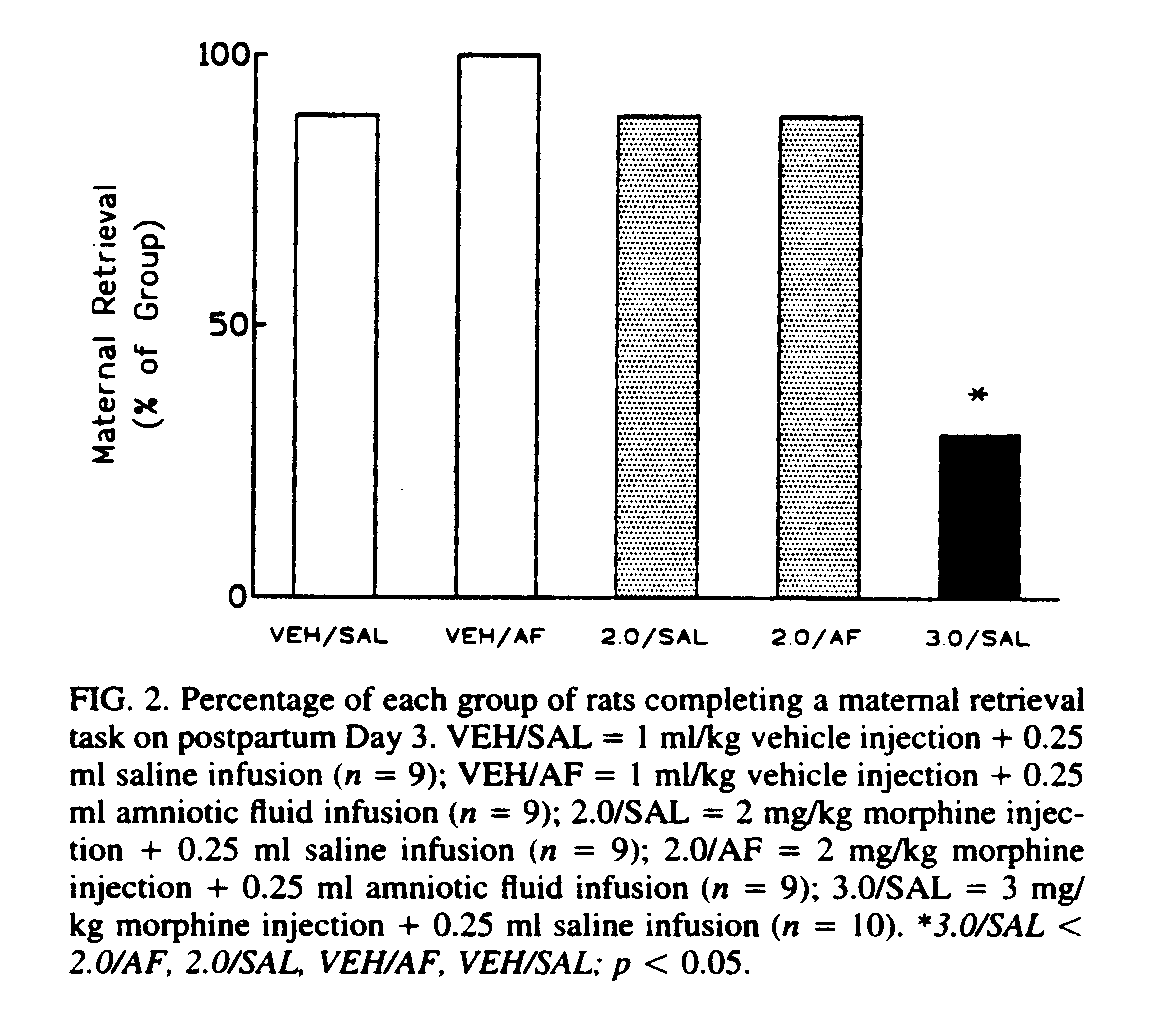
More recent research has focused on the differential effects of POEF administration on various opioid-mediated phenomena (e.g., hyperthermia, locomotor activation, antinociception, maternal retrieval). These differential effects have led us to speculate that either POEF differentially modifies different classes of opioid receptors, or has location-specific effects, or both.
In an attempt to determine the extent to which POEF is opioid-receptor specific, DiPirro has conducted a series of studies in which the effect of ingested placenta was tested on rats receiving an intraventricular injection of a specific opioid-receptor agonist [2, 3, 6]. The agonists used were DPDPE for delta-opioid receptors; DAGO for mu-opioid receptors; and U-62066 (spiradoline) for kappa-opioid receptors. Different doses of agonist were injected in a constant volume through indwelling intracerebroventricular cannulae, and pain thresholds were assessed at peak agonist effect, using a 52° C hotplate, after ingestion of 1g of placenta or meat control. No repeated measures were used.
DPDPE -- The effect of placenta (and presumably POEF)
ingestion on delta-opioid antinociception produced by an intracerebroventricular
injection of DPDPE is presented in the next figure (Fig. 3):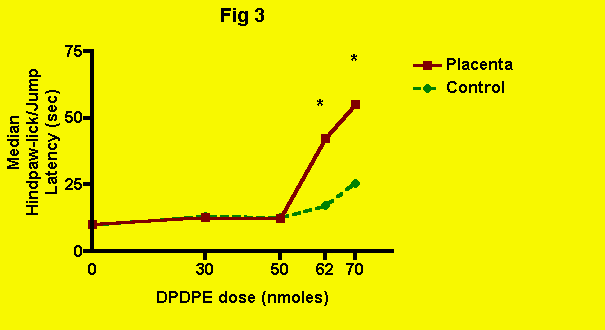 Clearly, placenta ingestion potentiated the effect of delta-opioid mediated
antinociception produced by DPDPE. Without placenta, DPDPE did not produce
a significant elevation of pain threshold at a dose of 62 nmoles. Even
at 70 nmoles, the increase in pain threshold was minor. With placenta ingestion,
however, DPDPE produced a 250% increase in pain threshold at a dose of
62 nmoles.
Clearly, placenta ingestion potentiated the effect of delta-opioid mediated
antinociception produced by DPDPE. Without placenta, DPDPE did not produce
a significant elevation of pain threshold at a dose of 62 nmoles. Even
at 70 nmoles, the increase in pain threshold was minor. With placenta ingestion,
however, DPDPE produced a 250% increase in pain threshold at a dose of
62 nmoles.
DAGO -- The effect of placenta (and presumably POEF) ingestion
on mu-opioid-mediated antinociception produced by the administration of
DAGO can be seen in the following figure (Fig. 4):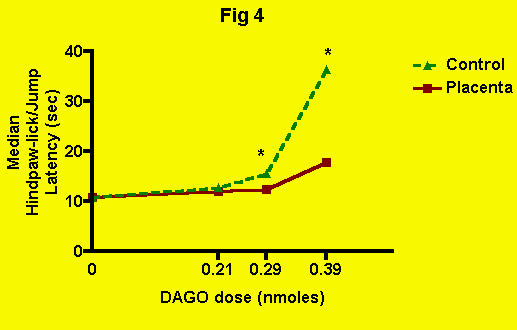 The antinociceptive effect of DAGO alone (along with ingestion of meat
control) appears at the 0.29-nmole dose. At 0.39 nmoles, the antinociception
is quite pronounced. However, in combination with placenta ingestion, a
significant attenuation of DAGO-induced antinociception becomes apparent
at the 0.29-nmole dose, and the elevation of pain threshold produced by
0.39 nmoles DAGO is all but eliminated. Therefore, placenta (and presumably,
therefore, POEF) ingestion, blocks antinociception mediated by mu-opioid
receptors.
The antinociceptive effect of DAGO alone (along with ingestion of meat
control) appears at the 0.29-nmole dose. At 0.39 nmoles, the antinociception
is quite pronounced. However, in combination with placenta ingestion, a
significant attenuation of DAGO-induced antinociception becomes apparent
at the 0.29-nmole dose, and the elevation of pain threshold produced by
0.39 nmoles DAGO is all but eliminated. Therefore, placenta (and presumably,
therefore, POEF) ingestion, blocks antinociception mediated by mu-opioid
receptors.
U-62066 -- The effect of placenta (and presumably POEF) ingestion on kappa-opioid-mediated antinociception produced by central administration of U-62066 (spiradoline) is illustrated in the following figure (Fig. 5):
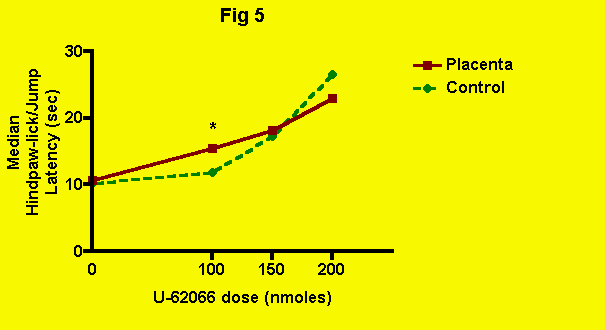
At the lowest dose of U-62 used, 100 nmoles, U-62 alone was clearly ineffective, whereas U-62 in conjunction with placenta ingestion, produced a significant elevation of pain threshold.
The effect of POEF on central opioid processes is almost certainly location
specific as well as receptor specific; very recent data indicate that antinociception
produced by morphine injected directly into the periaqueductal gray matter
is unaffected by placenta ingestion [4]. Opioid receptors are differentially
distributed, and clearly show different effects at different locations.
Bridges' work, for instance, has demonstrated that increased opioid levels
in the medial preoptic area interfere with maternal behavior [1, 10, 19].
In contrast, Thompson's recent work has shown that increasing the opioid
activity of the ventral tegmental area (by injecting morphine), which also
increases motivated behavior, facilitated the onset of maternal behavior
in inexperienced, nonpregnant rats. Conversely, blocking the effect of
morphine injection in the ventral tegmental area, by pretreating the rats
with the opiate antagonist naltrexone, blocked the facilitative effect
on maternal behavior of the intra-tegmental morphine injection. Furthermore,
interfering with the effect of endogenous opioids in the ventral
tegmental area at the end of parturition by intra-tegmental
injection of naltrexone methobromide, severely
inhibited the naturally-occurring onset of maternal behavior at that
time [24]. The following figure (Fig 6) shows the deleterious effect on
the rate of onset of maternal behavior, at parturition, of naltrexone methobromide
(quaternary naltrexone) injected into the ventral tegmental area.
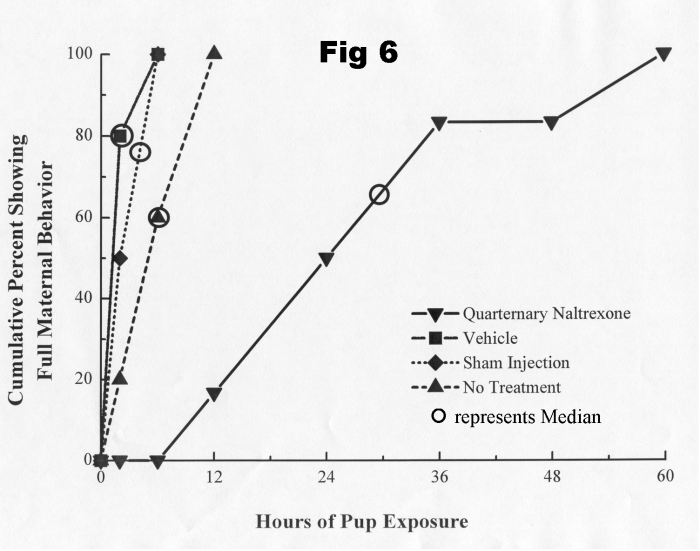
Although POEF ingestion can modify the effects of intra-tegmental morphine on locomotor behavior [24], we do not yet know if POEF ingestion potentiates the effect of intra-VTA morphine on maternal behavior; POEF, ingested in amniotic fluid, however, did not increase the effect of a subthreshold systemic dose of morphine on maternal retrieval to the point where it disrupted the retrieval of young by mother rats (see Fig. 2).
The mechanical stimulation and distention of the vaginal/cervical area during expulsion of the fetus is well above the minimum amount necessary to trigger pseudopregnancy in nonpregnant females experiencing elevated estrogen levels. Yet the parturient rat does not enter pseudopregnancy, but rather experiences a postpartum estrus and, if inseminated, a postpartum pregnancy. We hypothesized, based on some supportive pilot data, that afterbirth ingestion would decrease the likelihood that relatively intense vaginal/cervical stimulation would induce pseudopregnancy. We tested this hypothesis in a study [23] in which groups of nonpregnant rats received vaginal/cervical stimulation of pressures of either 75, 125, 175 or 225 g. The stimulation applied to the vaginal/cervical area, as expected [14], produced a dose-dependent level of antinociception (measured as lengthening of tail-flick latency or hot-water tail-withrawal latency). In addition, orogastric infusion of amniotic fluid, shortly before the application of vaginal/cervical stimulation, enhanced the level of antinociception produced by 125 g pressure. Interestingly, the 225-g level of stimulation induced pseudopregnancy in 44% of the rats receiving an orogastric infusion of saline, but only in 10% of the rats infused with amniotic fluid. A detailed series of follow-up studies on the effect of amniotic-fluid ingestion on pseudopregnancy induction is currently being conducted by Patricia Abbott.
Mechanical stimulation during delivery, resulting from expulsion and activity of the fetus, produces major activation of the pelvic, hypogastric [15], and pudendal nerves. This afferent information, including nociceptive, results in a central release of endogenous opioids. The opioids, in turn, orchestrate a variety of parturitional phenomena including an elevation of pain threshold, the onset of maternal caretaking behaviors, and even perhaps a reduction in the likelihood that this vaginal/cervical stimulation will induce pseudopregnancy, a condition that would eliminate the postpartum estrus. Paradoxically, a higher level of endogenous opioids, despite producing an apparently advantageous additional rise in pain threshold, would counteract some other beneficial effects of opioids, especially those relating to the emergence or performance of the complex of behaviors necessary for caretaking of the young. However, it is apparent that a substance is available in afterbirth material (POEF) that, when ingested by the mother, potentiates the antinociceptive actions of endogenous opioids (raising pain threshold above that level produced solely by the opioids), but not the actions that would, if increased, have a deleterious effect on maternal behavior. This differential effect on certain opioid functions is the result of selective action on some classes of opioid receptor (as well as some degree of location specificity). Mu-opioid receptor activity, largely associated with negative "side" effects of opioid activity, is suppressed by POEF, whereas kappa- and delta-opioid-mediated phenomena are enhanced. This is particularly interesting, and important, in light of recent studies showing that kappa-opioid analgesics (and to a lesser extent delta-opioid analgesics) are particularly effective in females [e.g., 7, 20].
REFERENCES
- Bridges, R.S.; & Grimm, C.T. (1982). Reversal of morphine disruption of maternal behavior by concurrent treatment with the opiate antagonist naloxone. Science 218:166-168.
- DiPirro, J.M. & Kristal, M.B. (1994). Analgesia produced by ICV injection of DPDPE in rats is enhanced by placenta ingestion. Society for Neuroscience Abstracts, 20, Part 1, 752.
- DiPirro, J.M. & Kristal, M.B. (1997). Placenta ingestion facilitates locomotion and antinociception induced by activation of delta-opioid receptors in the rat. Society for Neuroscience Abstracts, 23, Part 1, 680.
- DiPirro, J.M. & Kristal, M.B. (1998). The effect of placenta ingestion on antinociception induced by morphine in the periaqueductal gray of the rat. Society for Neuroscience Abstracts, 24, Part 1, 1132.
- DiPirro, J.M., Thompson, A.C., & Kristal, M.B. (1991). Amniotic-fluid ingestion enhances the central analgesic effect of morphine. Brain Research Bulletin 26: 851-855.
- DiPirro, J.M., Vanderwerf, T.M. & Kristal, M.B. (1996). The effect of placenta ingestion on kappa-opioid antinociception in rats. Society for Neuroscience Abstracts, 22, Part 2, 1365.
- Gear, R.W.; Miaskowski, C; Gordon, N.C.; Paul, S.M.; Heller, P.H.; Levine, J.D. (1996). Kappa-opioids produce significantly greater analgesia in women than in men. Nature Medicine 2: 1248-1250.
- Gintzler, A.R. (1990). Endorphin-mediated increase in pain threshold during pregnancy. Science 210: 193-195.
- Graber, G.C. & Kristal, M.B. (1977). Uterine distention facilitates the onset of maternal behavior in pseudopregnant but not in cycling rats. Physiology & Behavior 19: 133-137.
- Grimm, C.T.; Bridges, R.S. (1983). Opiate regulation of maternal behavior in the rat. Pharmacology Biochemistry & Behavior 19: 609-616.
- Keverne, E.B.; Levy, F.; Poindron, P. & Lindsay, D.R. (1983). Vaginal stimulation: An important determinant of maternal bonding in sheep. Science 219: 81-84.
- Kristal, M.B. (1991). Enhancement of opioid-mediated analgesia: A solution to the enigma of placentophagia. Neuroscience & Biobehavioral Reviews 15: 425-435.
- Kristal, M.B., Tarapacki, J.A. & Barton, D. (1990). Amniotic fluid ingestion enhances opioid-mediated but not nonopioid-mediated analgesia. Physiology & Behavior 47: 79-81.
- Kristal, M.B., Thompson, A.C., Heller, S.B., & Komisaruk, B.R. (1986). Placenta ingestion enhances analgesia produced by vaginal/cervical stimulation in rats. Physiology & Behavior 36: 1017-20.
- Peters, L.C., Kristal, M.B., & Komisaruk, B.R. (1987). Sensory innervation of the external and internal genitalia of he female rat. Brain Research 408: 199-204.
- Pinheiro Machado F° , L.C., Hurnik, J.F., & Burton, J.H. (1997). The effect of amniotic fluid on the nociception of cows. Physiology & Behavior 62: 1339-1344.
- Robinson, T.M., Abbott, P. & Kristal, M.B. (1995) Blockade of digestion by famotidine pretreatment does not interfere with the opioid-enhancing effect of ingested amniotic fluid. Physiology & Behavior 57: 261-263.
- Robinson-Vanderwerf, T.M., DiPirro, J.M., Caggiula, A.R. & Kristal, M.B. (1997). The analgesia-enhancing component of ingested amniotic fluid does not affect nicotine-induced antinociception in naltrexone-treated rats. Pharmacology Biochemistry & Behavior 58: 147-151.
- Rubin, B.S.; Bridges, R.S. (1984). Disruption of ongoing maternal responsiveness in rats by central administration of morphine sulfate. Brain Research 307: 91-97.
- Sander, H.W.; Portoghese, P.S.; Gintzler, A.R. (1988). Spinal kappa-opiate receptor involvement in the analgesia of pregnancy: Effects of intrathecal nor-binaltorphimine, a kappa-selective antagonist. Brain Research 474: 343-347.
- Tarapacki, J.A., Piech, M. & Kristal, M.B. (1995). Ingestion of amniotic fluid by postpartum rats enhances morphine analgesia without liability to maternal behavior. Physiology & Behavior 57:209-212.
- Tarapacki, J.A., Thompson, A.C. & Kristal, M.B. (1992). Gastric vagotomy blocks opioid-analgesia enhancement produced by placenta ingestion. Physiology & Behavior 52: 179-82.
- Thompson, A.C., Ferguson, E.J., Abbott, P., Doerr, J.C. & Kristal, M.B. (1991) Amniotic-fluid ingestion before vaginal/cervical stimulation produces a dose-dependent enhancement of analgesia and blocks pseudopregnancy. Physiology & Behavior 50: 11-15.
- Thompson, A.C. & Kristal, M.B. (1996). Opioid stimulation in the ventral tegmental area stimulates maternal behavior in rats. Brain Research 743: 184-201.
- Yeo, J.A.G. & Keverne, E.B. (1986). The importance of vaginal-cervical stimulation for maternal behaviour in the rat. Physiology & Behavior 37:23-26.
| Discussion Board| Previous Page| Your Symposium |